Анеуплоиды
Анеуплоиды - это организмы, у которых произошло увеличение или уменьшение числа отдельных хромосом в геноме. Анеуплоиды нельзя непосредственно использовать в производстве, но при создании рядов моносомиков или трисомиков для каждой хромосомы они могут оказаться полезными для замены или дополнения отдельных хромосом в уже выведенных сортах. В настоящее время ученым удалось создать ряды моносомиков и трисомиков следующих видов сельскохозяйственных растений:

Исчерпывающий анализ и литературные данные о создании, характеристике и использовании анеуплоидов растений дали Сирс и Куш.
Исследования моносомиков и трисомиков вскрывают роль отдельных хромосом в выраженности признаков и развитии организма как единого целого. Изучение моносомиков позволяет определять в хромосомах места расположения генов, ответственных за отдельные признаки. Если необходимо установить, в какой хромосоме данного сорта находится ген (или гены) устойчивости к листовой ржавчине, этот сорт скрещивают с другим сортом, имеющим моносомную серию. При доминантной устойчивости, определяемой одним единственным геном, в поколении F2 от скрещивания с любыми другими хромосомами (за исключением одной) можно ожидать появления 75% устойчивых и 25% неустойчивых растений. Согласно системе скрещивания, в которое включено только устойчивое растение, можно сделать заключение, что ген устойчивости находится в хромосоме растения-моносомика. Такое скрещивание можно представить следующим образом:

Если встает вопрос об определении положения рецессивных генов, то уже в F1 можно установить хромосомы, являющиеся носителями этих генов:

С помощью моносомных рядов в настоящее время удалось определить положение огромного числа генов в отдельных хромосомах. Например, установлено, что ген Lr 9, ответственный за устойчивость к листовой ржавчине пшеницы, расположен в длинном плече хромосомы 6 В (6 BL), а ген Hg, ответственный за опушение колосковых чешуй, - в коротком плече хромосомы 1А.
Замещение и добавление хромосом
Создание моносомных рядов позволяет проводить замещение отдельных хромосом одного сорта на хромосомы другого сорта. При скрещивании двух сортов едва ли удастся найти растения, несущие все хромосомы одного сорта и только одну хромосому того сорта, чьи признаки нужно передать первому. Наличие моносомных рядов делает это возможным. Точно так же можно передавать не весь набор хромосом, а отдельные хромосомы от других видов, хотя при этом нередко возникают огромные трудности. Эти новые и интересные возможности будут рассмотрены на ряде примеров.
Сирс хотел передать пшенице обыкновенной устойчивость к листовой ржавчине от Aegilops umbellulata. Так как эти два вида не поддавались скрещиванию, он взял в качестве посредника Tr. dicoccoides.

В ходе этого скрещивания было получено одно растение с 42 хромосомами пшеницы и одной хромосомой эгилопса, устойчивое к листовой ржавчине. Однако добавление хромосомы одновременно привело к появлению таких нежелательных признаков, как ломкость основной ости колоса, стерильность пыльцы и т.д. Поэтому потребовалось отделить часть хромосомы эгилопса, которая является носителем устойчивости к ржавчине, от части - носителя отрицательных признаков. Потомство этой формы Сирс обработал, рентгеновскими лучами и облученную пыльцу использовал для опыления. Из 6091 растения получили 40 растений с транслоцированными участками хромосомы эгилопса на хромосому 6В пшеницы обыкновенной, в то же время оставшаяся часть, или фрагмент хромосомы эгилопса, была утрачена и с ней исчезли отрицательные признаки. Так получили новый сорт Transfer, служащий источником устойчивости к листовой и стеблевой ржавчине.
Аналогичных результатов достиг Дрисколл при скрещивании пшеницы и ржи, а также при облучении растений, имевших все хромосомы пшеницы и одну пару хромосом ржи. В результате была получена линия Transec, содержащая один сегмент хромосомы ржи, транслоцированный на хромосому 4А и несущий устойчивость к листовой ржавчине и мучнистой росе.
Добавление или передачу хромосомы от одного вида другому уже применяли в ряде случаев. Создание линии замещения путем переноса хромосомы ржи в хромосомный набор пшеницы одними из первых провели Эванс и Дженкинс, а также другие исследователи. Этого можно достичь методом, который схематично изображен ниже:

Линии замещения, полученные у ржи, овса, пшеницы, табака, не дали пригодных товарных сортов, так как генотипы с одной и особенно с большим числом пар хромосом от чужих сортов нестабильны и такие хромосомы часто утрачиваются.
Замещение хромосом - гораздо более перспективный метод в селекции растений. Если в какой-либо продуктивный, но неустойчивый к листовой ржавчине сорт нужно перенести ген Lr9, то можно заменить его хромосому 6В на хромосому 6В линии Transfer:

Следовательно, в поколении F1 скрещивание донора-дисомика (Transfer) с реципиентом-моносомиком (Panonija), имеющим хромосому 6В (носитель восприимчивости к листовой ржавчине), дает растение, которое имеет 20 бивалентов (от каждого родителя по 20 хромосом) и один унивалент с 6В от сорта-донора. Так как цель заключается в получении всех признаков от реципиента, или рекуррентного родителя, а от сорта-донора - только одной пары хромосом (6B), Нужно вначале провести не менее 4-6 возвратных скрещиваний с моно-6B Panonija, а затем - принудительное самоопыление. Это даст растения-дисомики, которые будут содержать в основном все пары хромосом от рекуррентного родителя и хромосому 6В с геном устойчивости к ржавчине от донора.
Таков пример внутривидового замещения, т.е. замещения между двумя сортами внутри одного вида. Однако можно проводить замещение хромосом между различными видами и даже родами. Прогресс, достигнутый в этом направлении у пшеницы, позволил Рилею и Окамото установить, что конъюгация гомологичных хромосом контролируется доминантным геном, расположенным в длинном плече хромосомы 5В. Следовательно, при конъюгации хромосомы 5В (растение-нуллисомик по длинному плечу 5В) наступает конъюгация гомеологических хромосом, и таким образом можно провести замещение. Рилей передал пшенице обыкновенной устойчивость к Puccinia striiformis от Aegilops comosa следующим образом:

Так было получено растение с 42 хромосомами пшеницы и одной хромосомой 2М эгилопса. Поскольку линия замещения была нестабильной, а в дальнейших поколениях произошла бы утрата хромосомы эгилопса, растение скрестили с видом Ае. speltoides, у которого произошла конъюгация 5В хромосом:
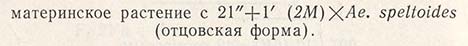
После нескольких возвратных скрещиваний с Chinese Spring и отбора на устойчивость к ржавчине были получены:

На стадии завершения этого процесса гибридизации получен сорт Compair, имеющий 20 бивалентов от пшеницы обыкновенной и одну хромосому 2М от Ае. comosa, конъюгирующую с одной хромосомой 2D сорта Chinese Spring. Растения этого сорта имеют правильно протекающий мейоз и могут быть использованы для передачи признака устойчивости к P. striiformis без дополнительных затруднений, связанных с межвидовой гибридизацией.
Выше были приведены только некоторые из новейших примеров, показывающих, что в отыскании путей и методов получения организмов с новыми признаками существуют огромные возможности. Лоу и Уорланду в опытах с несколькими сортами пшеницы, неустойчивыми к P. striiformis, удалось провести замещение хромосомы 5Bs/7Bs от сорта Bersee, устойчивого к желтой ржавчине ржи.
В некоторых случаях можно провести замещение данной хромосомы хромосомой другого вида, но только если она генетически сходна с замещаемой хромосомой реципиента. Так, сорта пшеницы Orlando, Zorba, Weique, Neuzucht, Clement содержат хромосому 1R от ржи вместо своей хромосомы 1В. Кроме того, известные сорта Аврора и Кавказ, в родословные которых входит сорт Neuzucht, содержат транслоцированный сегмент хромосомы 1R в хромосоме 1В.
Таким образом, анеуплоидия стала одним из современных методов селекции растений. Особая ценность анеуплоидии заключается в том, что она позволяет создавать новые сорта, имеющие только одну или две хромосомы от тех сортов, чьи признаки хотят совместить со всеми остальными хромосомами сорта-реципиента. Это особенно важно при межвидовой гибридизации, так как свободное комбинирование хромосом от двух видов в большинстве случаев очень затруднено: совмещение положительных признаков связано с появлением ряда отрицательных, которые сводят почти на нет позитивные стороны межвидовой гибридизации.
Следует подчеркнуть, что анеуплоидия - это довольно долгий путь получения нового материала и особенно новых сортов для производства, и поэтому она главным образом служит одной из ступеней в селекции растений с применением гибридизации.