Гаплоиды
Гаплоиды - это организмы, имеющие в соматических клетках такое же число хромосом (n), как и в гаметах своего вида. Одним из первых обнаруженных и изученных примеров гаплоидов у растений является Datura stramonium; позже они были найдены у табака, томатов, кукурузы, пшеницы и других видов.
Гаплоиды чаще всего возникают из неоплодотворенной яйцеклетки (гиногенез), из других клеток гаметофита (апогамия) или из мужских гамет (андрогенез) и часто - у полиэмбрионов. Поскольку гаплоиды могут иметь различные геномные формулы в зависимости от уровня плоидности вида, от которого они произошли, необходимо прежде всего познакомиться с названиями диплоидов и гаплоидов (табл. 18.3). Частота возникновения гаплоидов в природе весьма низка: около 1 на 1000 при партеногенезе и около 0,1 на 1000 при андрогенезе.
В большинстве случаев гаплоидные растения характеризуются меньшей высотой стебля, слабым развитием, позднеспелостью; из-за наличия унивалентов вместо нормальных хромосом они в основном полностью стерильны. Поэтому гаплоиды сохраняются только у видов, размножающихся вегетативно, а у остальных видов элиминируют.
Если у гаплоидных растений не наступает редукционного деления в макро- и микроспорогенезе или число их хромосом удваивается под влиянием обработок колхицином, то растения становятся полностью фертильными и (что самое интересное) полностью гомозиготными по всем аллельным генам, которые они содержат. Понимание этого открывает возможности для использования гаплоидии в генетических исследованиях и селекции растений. Так, исследователи пришли к методам индуцирования гаплоидов in vitro и in vivo, чтобы повысить частоту их появления у различных видов растений. Этому вопросу были посвящены специальный симпозиум и публикация под редакцией Каша.
Культура пыльников стала самым успешным методом in vitro для индуцирования гаплоидов из пыльцы. К настоящему времени они получены приблизительно у 60 видов, из которых 90% приходится на семейства Solanacea и Gramineae. Однако определение наиболее благоприятного состава питательной среды, фаз развития пыльцевого зерна, влияния температуры и освещенности связано с трудностями, и производство гаплоидов из пыльцы стало обычным методом только у табака и Datura.
Отсюда возникла необходимость поиска метода индуцирования гаплоидов in vivo. У целого ряда видов растений (Solarium, Medicago, Trifolium, Raphanus, Brassica и др.) развитие зародыша происходит без оплодотворения яйцеклетки. Следовательно, изменяя режим опыления, проводя обработку химическими веществами или применяя воздействие чужеродной цитоплазмы после межвидовых скрещиваний, можно повысить частоту выхода гаплоидов.

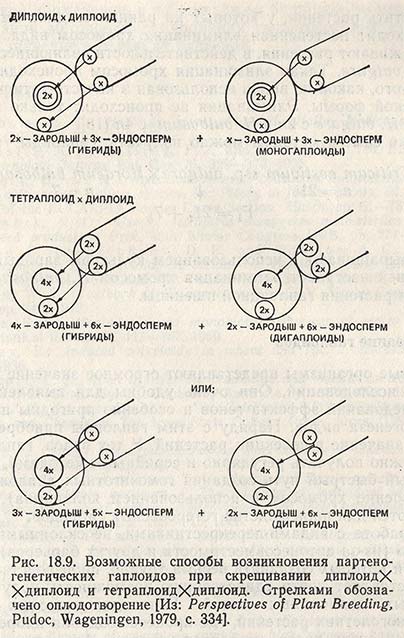
При скрещивании диплоида с диплоидом у перечисленных видов растений наряду с возникновением нормальных растений (2х зародыш и Зx эндосперм) в некоторых случаях оплодотворение яйцеклетки одним чужеродным генеративным ядром спермия отсутствует, и вместо него происходит оплодотворение вторичного зародышевого мешка (2х) и второго генеративного ядра спермия (x). В результате развивается моногаплоидный организм, имеющий в зародыше гаплоидное число хромосом (x) и триплоидный эндосперм (3x) (рис. 18.9).
Точно так же при скрещивании тетраплоида с диплоидом в результате партенокарпии развиваются дигаплоиды с зародышем 2х и эндоспермом 6х вместо 4х + 6х (рис. 18.9). Например, у люцерны, являющейся автотетраплоидом, в качестве опылителя для индуцирования дигаплоидов используют диплоидный вид Medicago falcata; у земляники, являющейся октоплоидом, в качестве опылителя берут тетраплоид Potentilla anseritia; у Solarium tuberosum опылителем может служить диплоидный вид S. phureja.
Встречаются и случаи так называемой семигамии, когда одна генеративная клетка пыльцевого зерна проникает в зародышевый мешок, но слияния с яйцеклеткой не происходит, а развивается химерное гаплоидное растение.
Для обнаружения гаплоида среди семян необходимо воспользоваться каким-то маркером, выделяющимся по окраске зародыша семени. Так, у кукурузы используют доминантный ген антоциановой окраски зародыша (Чейз, 1950), а у Solarium ssp. - ген окраски чашечки цветка. Необходимо лишь, чтобы носителем этого доминантного гена было отцовское растение. У нормального гибридного растения F1 все семена будут выделяться окраской зародыша, а гаплоиды останутся бесцветными, как у материнского растения.
К особым методам получения гаплоидов in vivo относится метод селективной элиминации хромосом. Между некоторыми видами может произойти скрещивание, в результате которого элиминируют хромосомы одного вида и остается только гаплоидное число хромосом другого вида.
При скрещивании Hordeum vulgare с дикорастущим Hordeum butbosum получают семена с очень недоразвитым эндоспермом. Если такие семена использовать для культуры зародыша, то можно вырастить растения, у которых на ранних стадиях эмбриогенеза происходит постепенная элиминация хромосом вида H. bulbosum и выживают растения, в действительности являющиеся гаплоидами H. vulgare. Такая элиминация хромосом происходит независимо от того, какой из видов использован в качестве отцовской или материнской формы. Элиминация не происходит только при скрещивании H. vulgare с 2n и H. bulbosum с 4n. Тем же самым методом можно получать гаплоиды пшеницы:
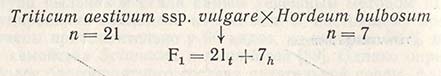
Семена выращивают с использованием культуры зародыша, через 10-14 дней наступает элиминация хромосом H. bulbosum и формируются растения гаплоидной пшеницы.
Использование гаплоидов
Гаплоидные организмы представляют огромное значение для генетических исследований. Они очень удобны для выявления мутаций, исследований эффекта генов и особенно пригодны при изучении филогенеза видов. Наряду с этим гаплоиды приобретают все большее значение в селекции растений. У тех видов, гаплоиды которых можно получать регулярно и серийными методами, открывается самый быстрый путь создания гомозиготных диплоидных линий (удвоение хромосом с использованием колхицина), которые применяются для производства гетерозисных гибридов.
При работе с видами-перекрестниками, не склонными к самоопылению (из-за автонесовместимости и других барьеров), инбредные линии которых создавать весьма трудно, производство гаплоидов представляет почти единственную возможность получения гомозиготных диплоидных линий. Гаплоидия у лесных пород и других многолетних растений, где процесс создания инбредных линий длится очень долго, позволяет ускорить формирование гомозиготных линий, используемых для получения гибридов.
В будущем гаплоиды могут найти применение при создании линий с цитоплазматической мужской стерильностью, которые служат для выведения гетерозисных гибридов многих видов (кукуруза, подсолнечник, рис и др.). Скрещиванием одной цмс-линии с гомозиготной диплоидной линией, полученной андрогенетическим путем (из пыльцы) от цмс-моногаплоида, у которого колхицином удвоили число хромосом, получают гомозиготную диплоидную цмс-линию.
Производство дигаплоидов картофеля методически настолько отработано, что позволяет широко практиковать перенос отдельных генов устойчивости к различным патогенным организмам. Таким образом, создание гаплоидных растений находит все более широкое применение, и в ближайшем будущем, по мере усовершенствования методов индуцирования гаплоидии, оно приобретет огромное значение в селекции растений.