Рекомбинации генов
Если родители различаются только двумя парами аллельных генов, то при отсутствии сцепления между этими генами и полном доминировании одного гена над другим (в результате расхождения хромосом с половыми и независимыми репродуктивными материнскими и отцовскими гаметами поколения F1 в F2) развиваются два новых гомозиготных генотипа, несущих новые комбинации генов. В этом случае можно говорить о рекомбинации генов у обоих родителей, схематично показанной ниже:

При первом скрещивании родителей, различающихся тремя парами аллельных генов, в F2 развивается шесть новых генотипов, отличающихся от родителей:

Если растения отобраны для скрещивания по 4, 5, 10 и более парам аллельных генов (n), что происходит чаще всего, то вследствие гетерозиготности организмов поколение F1 дает огромное число (2n) различных гамет. В условиях свободного размножения это приводит к формированию в F2 очень большого числа (3n) разных генотипов и 2n разных фенотипов. Таким образом, возникает новая, огромная по своим масштабам генетическая изменчивость, которая служит для селекционера источником для отбора новых комбинаций признаков, т.е. для создания новых линий и сортов.
Рекомбинация генов с доминантным, относительно рецессивным эффектом от двух и более родителей в одном генотипе обусловливает огромную генетическую изменчивость и приводит к созданию сортов с новыми свойствами. Однако это всего лишь комбинация существующих родительских генов. Если гены не сцеплены, новые комбинации признаков или в широком смысле рекомбинации генов можно предусмотреть по частоте и времени их возникновения (поколениям). Если же гены сцеплены, рекомбинацию генов как результат кроссинговера предвидеть невозможно, так же как нельзя предусмотреть частоту накопления генов, не установив предварительно удаленность соответствующих локусов в хромосомах.
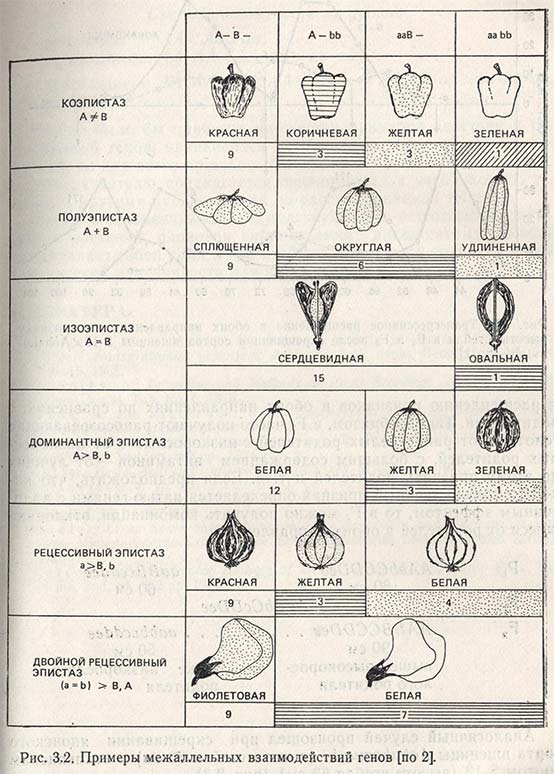
Между тем, учитывая различные типы проявления действия генов и их межаллельного взаимодействия, можно получать генотипы с такими свойствами и признаками, которых не было ни у одного родителя. Это случается, когда два или более разных гена обусловливают выраженность одного признака, но имеют при этом различные эффекты действия. Речь идет о коэпистатическом, полуэпистатическом, изоэпистатическом и гипостатическом эффекте, а также и об ингибиторном и комплементарном действии генов. Это и есть источники новой генетической изменчивости, некоторые варианты которых показаны на рисунке 3.2. Таким путем получают сорта с окрашенными цветками или цветками, лишенными окраски, сорта с разной формой плодов, различными химическими свойствами зерна и т.д.
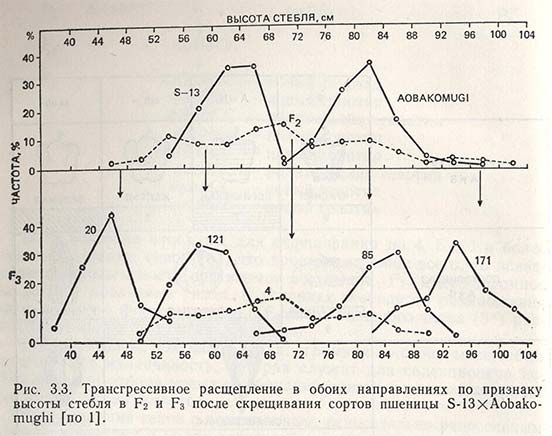
Кроме того, необходимо выделить трансгрессию генов, являющуюся источником новой генетической изменчивости, которая ведет к расщеплению признаков в обоих направлениях по сравнению с родителями. Таким образом, в F2 часто получают раносозревающие генотипы от раннеспелых родителей, низкорослые - от низкорослых родителей, с большим содержанием витаминов - от лучших по этому признаку родителей и т.д. Если предположить, что какой-то количественный признак определяется пятью генами с аддитивным эффектом, то в F2 можно получить комбинации, отклоняющиеся от родителей в обоих направлениях:

Аналогичный случай произошел при скрещивании японского сорта пшеницы Aobakomughi (высота стебля 83 см) с итальянским сортом S-13 (высота стебля 63 см) (рис. 3.3).
Приведенные примеры касались эффекта и взаимодействия генов, которые приводят к новым рекомбинациям признаков на диплоидном уровне. Размах генетической изменчивости возрастет еще больше, если взаимодействие происходит на разных уровнях плоидности, а именно:

Можно было бы привести множество примеров действия и рекомбинаций генов, являющихся источниками генетической изменчивости, но для этого пришлось бы выйти за рамки данной книги. Поэтому читателю предлагается ознакомиться с учебником по генетике и другими публикациями по рассматриваемой теме.
Наравне с изменчивостью, существующей в естественных популяциях растений, плановая гибридизация дивергентных особей представляет собой один из богатейших источников создания новой генетической изменчивости для нужд селекции.