Внутривидовая гибридизация
Закономерности в механизме наследования признаков, открытые Г. Менделем (1865), а позднее подтвержденные и развитые дальше плеядой генетиков и селекционеров, обусловили превращение селекции живых организмов из эмпирики в науку. Осмысление того, что путем скрещивания или гибридизации родительских форм с различными признаками можно получить потомство, обладающее признаками одного и другого родителя, привело к возникновению плановой гибридизации, прогнозированию рекомбинаций генов и программированному созданию сортов с заданными признаками.
В дикорастущих популяциях перекрестно-размножаемых организмов, в силу того что свободное скрещивание постоянно ведет к гибридизации разных генотипов, существует огромный запас генетической изменчивости. Путем гибридизации, которую проводит человек, особенно у автогамных организмов, возникает и может возникать дальше генетическая изменчивость по масштабам больше той, что формируется в дикорастущих популяциях. Уже проведены скрещивания многих разновидностей и видов, которые вследствие имеющих место в природе явлений несовместимости, механических барьеров или географических изоляций никогда не произошли бы естественным путем.
В главе 3 показано, какие рекомбинации генов можно получить путем гибридизации генетически различных родителей. Гибридизация позволяет создать генофонд изменчивости, который не может возникнуть каким-либо другим способом. Наряду с этим гибридизация - это неизбежный этап всех других распространенных приемов и методов селекции растений. Поэтому, как и в прошлом, она останется основным средством и направлением в создании новых сортов растений и пород животных.
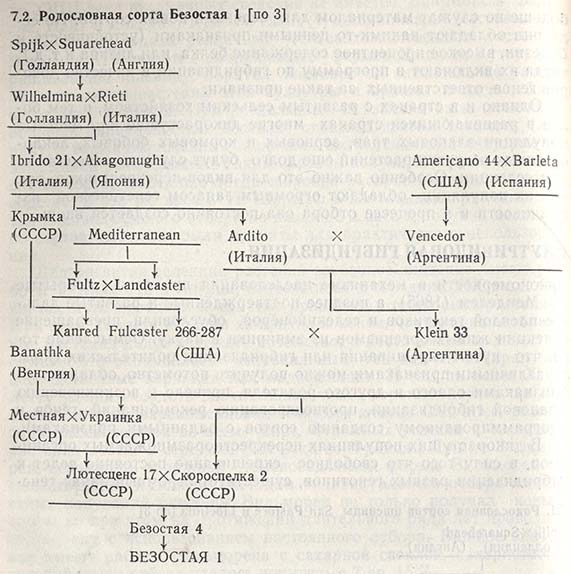
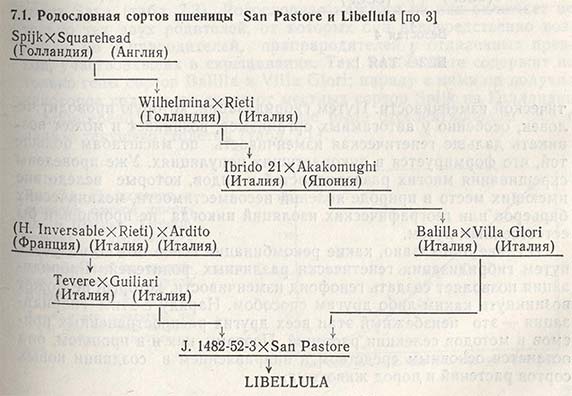
Учитывая это, почти в каждой главе данной книги говорится о различных характерных признаках гибридизации, здесь же ее отличительные черты и значение для селекции растений больше излагаться не будут. И тем не менее в качестве примера можно привести родословные итальянского сорта пшеницы San Pastore (табл. 7.1), советского сорта Безостая 1 (табл. 7.2) и югославского сорта Sava (табл. 7.3). Родословная каждого из них включает не только тех двух родителей, от которых они непосредственно возникли, но и прародителей, прапрародителей и отдаленных предков, участвовавших в скрещивании. Так, San Pastore содержит не только гены сортов Balilla и Villa Glori; наряду с ними он получил некоторое количество генов от местных сортов Spijk из Голландии и Squarehead из Англии, которые были отселектированы еще в начале текущего столетия (табл. 7.1).
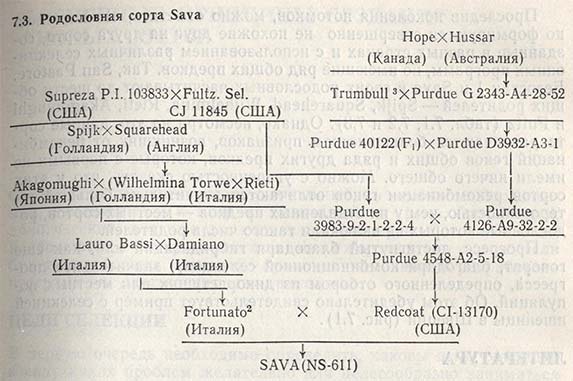
Проследив поколения потомков, можно отметить, что постепенно формируются совершенно не похожие друг на друга сорта, созданные в разных странах и с использованием различных селекционных программ, но имеющие ряд общих предков. Так, San Pastore, Безостая 1 и Sava в своих родословных насчитывают до шести общих родителей - Spijk, Squarehead, Wilhelmina, Rieti, Akagomughi и Fultz (табл. 7.1, 7.2 и 7.3). Однако, несмотря на это, данные сорта различаются большинством признаков, возникших от рекомбинаций генов общих и ряда других предков, которые с первыми не имели ничего общего. Можно с уверенностью сказать, что у этих сортов рекомбинации генов отличаются значительно большей гетерогенностью, чем у их отдаленных предков - местных сортов, родословные которых не включали такого числа родителей.
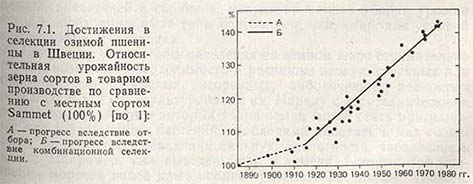
Прогресс, достигнутый благодаря гибридизации или, как еще говорят, благодаря комбинационной селекции, значительнее прогресса, определенного отбором из дикорастущих или местных популяций. Об этом убедительно свидетельствует пример с селекцией пшеницы в Швеции (рис. 7.1).