Фитогеографическая дифференциация как основа селекции растений
Значение селекции растений для развития общества лучше всего отражает высказывание великого генетика и селекционера Н. И. Вавилова: «Селекция растений - это эволюция руками человека». Познав закономерности наследования и применяя различные методы гибридизации и отбора, человек в течение десятилетий и столетий создал ряд новых разновидностей, на образование которых в природе потребовались бы тысячи и миллионы лет или которые вообще могли бы не возникнуть. Как установил Дарвин, в основе эволюции живых организмов лежит их изменчивость. Успех селекции растений также определяется внутри- и межвидовой изменчивостью, т.е. той, которая постоянно существует в природе, а также изменчивостью, созданной трудом человека.
Центры происхождения культурных растений
Генетическая дивергентность культурных видов растений и ее значение для дальнейшей селекционной работы особенно подробно показаны в исследованиях Н. И. Вавилова и его соратников. С 1923 по 1931 г. Вавилов организовал несколько научных экспедиций в более чем 60 стран мира. В этих экспедициях было собрано свыше 300 тыс. образцов культурных растений и их диких сородичей.
Собранный материал в течение нескольких лет изучали во Всесоюзном институте растениеводства в Ленинграде, многочисленных зональных институтах и на опытных станциях СССР. Проведенные исследования позволили получить ценные данные о размещении ряда культурных видов, внутривидовом разнообразии, изменчивости отдельных признаков, частоте проявления и распределении генов, контролирующих отдельные признаки, возникновении признаков под влиянием определенных факторов внешней среды и т. д. На основе этих исследований Вавилов (1935) установил восемь центров происхождения культурных растений на земном шаре (рис. 2.1). Ниже приведены лишь основные характеристики этих центров, подробности читатель может найти в капитальном труде H. И. Вавилова «Теоретические основы селекции растений» (1935).
1. Китайский центр. Китайский центр - самый древний самостоятельный очаг мирового земледелия. Сформировался он раньше всех других центров, и отсюда ведут свое происхождение многие культурные растения. Этот центр включает горные и прилегающие низменные районы Центрального и Западного Китая. Важнейшие эндемические растения китайского центра - просо, гречиха, соя, огромное число бобовых; здесь же сосредоточено многообразие форм пайзы и чумизы. Китай занимает первое место в мире по богатству видового и родового состава груши, яблони, косточковых плодовых (персика и др.) и многих видов цитрусовых. В большинстве случаев эти виды представлены здесь множеством ботанических разновидностей и генотипов.
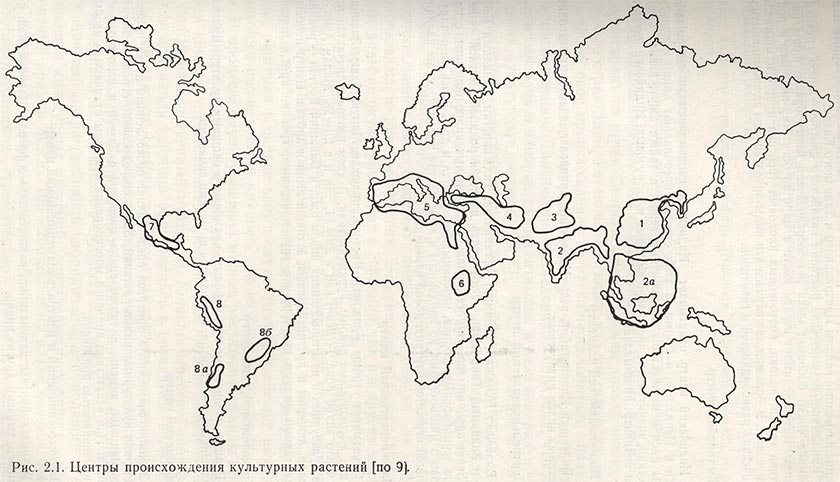
2. Индийский центр. Индийский центр охватывает Индию, за исключением ее северо-западной части и Пенджаба, и Бирму. В этих странах преобладают дикие и культурные виды риса с множеством доминантных генов, а также джут, кенаф, сахарный тростник, значительное число зернобобовых и цитрусовых. Индийский центр имеет один подцентр: индо-малайский.
2а. Индо-малайский подцентр. Включает Индокитай и весь Малайский архипелаг, здесь родина банана и некоторых видов цитрусовых.
3. Центральноазиатский центр. Включает Пенджаб, Кашмир, Афганистан, Таджикистан, Западный Узбекистан и Западный Тянь-Шань. Этот центр является родиной мягкой пшеницы (Triticum aestivum ssp. vulgare), ряда гексаплоидных видов пшеницы, основных видов зернобобовых (гороха, чечевицы, бобов, нута и др.). Здесь не обнаружено видового многообразия, но найденным в этом центре видам присуще наличие постоянной огромной внутривидовой изменчивости.
4. Ближневосточный центр. Охватывает страны Ближнего Востока, включая Малую Азию, Закавказье, Иран и высокогорные районы Туркменистана. В этом центре встречается огромное число видов рода Triticum как с 2n = 14, так и с 2n = 28. Малая Азия - родина культурных и некоторых диких видов ржи, а Ближний Восток - родина винограда, люцерны и других бобовых, а также дыни и арбуза. Центр необычайно богат такими видами культур, как груша, черешня, грецкий орех, миндаль и другие.
5. Средиземноморский центр. Родина маслины, многих бобовых, кормовых и овощных культур. Виды, происходящие из этого центра, отличаются крупными семенами.
6. Эфиопский центр. Эфиопия совместно с Эритреей представляет обособленный центр происхождения ряда культурных видов растений. Здесь сосредоточено огромное богатство разновидностей ячменя, а также видов пшеницы с 2п=28 и некоторых видов бобовых. При отсутствии видового многообразия все виды отличаются необычайной изменчивостью признаков.
7. Южномексиканский и центральноамериканский центр. Родина кукурузы, хлопчатника, какао, а также фасоли обыкновенной, перца овощного, тыквы и других видов.
8. Южноамериканский центр. В этом центре, включающем Перу, Эквадор и Боливию, сосредоточено большое число видов картофеля (Solatium ssp., 2n = 24, 36 и 60). Является также подиной томатов и табака. Южноамериканский центр имеет два подцентра: чилеанский и бразильско-парагвайский.
8а. Чилеанский подцентр. Этот подцентр - родина картофеля (Solatium tuberosum, 2n = 48) и ряда других видов.
8б. Бразильско-парагвайский подцентр. Этот подцентр - родина арахиса, маниока, ананаса и других видов.
Среди приблизительно 640 важнейших культурных видов более 500 происходят из стран Старого Света, причем только около 400 видов - из стран Азии. Установлено также, что значительное число видов ведет свое происхождение из нескольких центров, поэтому сейчас предполагают существование первичных и вторичных центров происхождения. Дальнейшие исследования показали, что правильнее говорить о центрах дивергентности, а не о центрах происхождения культурных растений, так как именно в местах происхождения культурных видов обычно обнаруживают их диких предшественников. По данным Вавилова, центры происхождения некоторых культур расположены довольно далеко от тех территорий, где сосредоточены их дикие сородичи. Так, в Эфиопском центре ячмень, пшеница, горох, лен и другие растения выделяются весьма значительной изменчивостью признаков, однако их дикие сородичи здесь вообще не найдены. Это свидетельствует о том, что данные виды не окультурены как в Эфиопии, так и в других местах. В настоящее время получены доказательства того, что эти виды были окультурены на Среднем Востоке во время земледельческой революции в период неолита и уже только после этого, вероятно, при переселении хамитов, попали в Эфиопию.
Харлан (1951, 1970) подчеркивает, что известные сегодня культурные растения в первое время окультуривания внешне выглядели намного иначе и были крупнее, чем их ближайшие дикие сородичи. В процессе окультуривания многие растительные виды по мере распространения за пределы своего центра происхождения вступали в контакт с родственными им дикими видами и получали от них новый наследственный материал. Хорошо известно, например, что гексаплоидная пшеница образовалась в результате включения зародышей плазмы Aegilops squarosa (интрагрессивная гибридизация) в тетраплоидные пшеницы, после чего уже последние были введены в культуру. Аналогично проходила передача наследственного материала от Tripsacum в кукурузу Zea mays.
О значении интрагрессивной гибридизации для эволюции и возникновения культурных растений свидетельствует один почти современный пример. Когда 100-200 лет тому назад человек осваивал Северную Америку и менял ее облик, однолетний подсолнечник Helianthus annuus стремительно распространялся и скрещивался при этом с некоторыми другими видами Helianthus. Гены различных видов, проникавшие в разные районы, способствовали тому, что Н. annuus адаптировался в различных экологических условиях и занял огромный ареал.
Поэтому Харлан (1970) считает, что происхождение культурных растений носит характер рассеивания во времени и в пространстве, что проблема «центра происхождения» никогда не была решена окончательно и правильнее было бы говорить о районах, где находился центр начала окультуривания, и о центрах дивергентности как о двух разных понятиях. Сам Вавилов пользовался термином «центры генетической дивергентности», но при классификации остановился на понятии «центры происхождения».
Для селекции растений особенно важны исследования Вавилова о генцентрах, т.е. о накоплении и распределении отдельных генов (окраски, формы, устойчивости к патогенам и т.д.) в определенных географических районах, что сегодня в основном необходимо при обнаружении источников генов.
Закон гомологических рядов в наследственной изменчивости признаков
Еще Дарвин, изучавший эволюцию разных видов растений, подметил существование параллелизма при варьировании морфологических признаков как внутри отдельного вида, так и между различными видами. Вавилов пришел к заключению, что при внутри- и межвидовом варьировании морфологических признаков речь идет обычно о проявлении повторяющегося сходства, определенной гомологии между рядами морфологических признаков. Например, в группе пшениц с 2n = 42 (Triticum ssp.) встречается несколько видов, ведущих происхождение из Центральноазиатского центра. Группа пшениц с 2n = 28 происходит из Эфиопии, Закавказья и Средиземноморья, а группа видов с 2n = 14 распространена в Малой Азии. Все эти виды географически удалены друг от друга и различаются многими морфологическими признаками, однако у них все же есть ряд общих признаков. Каждый из упомянутых здесь видов имеет разновидности: остистые и безостые, с темной, коричневой и белой окраской колоса, озимые и яровые, раннеспелые и позднеспелые, устойчивые и восприимчивые к болезням и т.д.
При сопоставлении двух относящихся к разным родам видов, например пшеницы и ржи, отчетливо видно, что как в одном, так и в другом роде встречаются озимые и яровые, раннеспелые и позднеспелые, мелкосемянные и крупносемянные разновидности и т.д. У яблони и персика, принадлежащих к разным родам, также проявляется сходство по ряду морфологических признаков. Гомологичную изменчивость можно обнаружить и между различными семействами, но, естественно, в меньшей степени, чем между родами или видами одного и того же рода.
Вавилов (1935) подчеркивал: «Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе расположены виды, тем полнее сходство в рядах их изменчивости». Эта закономерность показывает, что отдельный признак, отсутствующий в настоящий момент у одного вида, присущ другому виду, и появления этого признака в природе под влиянием спонтанных генных мутаций, взаимодействия генов при гибридизации или индуцирования мутаций можно ожидать и у третьего вида.
Так, внутри вида Triticum turgidum известны разновидности с ветвистым колосом, однако в природе у Tr. aestivum ssp. vulgare такие формы пока не найдены. За последние десятилетия от скрещивания ветвистых форм Тг. turgidumv.Tr. Vavilovi получены формы пшеницы обыкновенной с ветвистым колосом. У ячменя не обнаружено форм с безлигульными листьями, но они существуют у пшеницы и ржи; применение рентгеновских лучей позволило получить безлигульные мутанты ячменя.
Следовательно, знание гомологических рядов в наследственной изменчивости признаков, существующих внутри вида, между видами и родами, имеет огромное значение для создания сортов растений с новыми признаками.
Опасность ослабления генетической изменчивости
Знание центров генетической дивергентности позволяет использовать необходимые в селекционной работе гены, контролирующие отдельные признаки. Сегодня этот вопрос особенно актуален, так как по мере развития растениеводства и модернизации сельского хозяйства большинство местных популяций утрачено или сведено до малого числа разновидностей. В условиях интенсивного сельскохозяйственного производства, желая лучше использовать минеральные удобрения и механизацию, а также пытаясь снизить конкуренцию между растениями и повысить отдачу с единицы посевной площади, в настоящее время широко практикуют монокультуру. В результате сорта стали в значительной степени гомогенными, а уряда культур даже представляют чистые линии. Это стало особенно очевидным, когда высокопродуктивные мексиканские карликовые сорта пшеницы были интродуцированы в Индию, Пакистан и другие азиатские и африканские страны и высеяны на площадях в миллионы гектаров. И тем не менее в производстве ослабление изменчивости не привело к каким-либо катастрофам, скорее наоборот, удалось значительно повысить урожайность и улучшить технологию возделывания культур. Однако у некоторых экологов и генетиков это вызвало немалую тревогу: они выражают опасение, что в новых, генетически выравненных сортах масштабы генетической изменчивости постоянно сокращаются и это может привести к вспышкам эпифитотий, снижению продуктивности, а далее и к голоду.
Причины беспокойства, конечно, существуют, поскольку в некоторых случаях ослабление генетической изменчивости приводило к эпифитотиям, сопровождавшимся резкими снижениями продуктивности. Чанг (1979) приводит случаи:
- голода в Ирландии в 1840 г. из-за эпифитотии фитофтороза (Phytophtora infestans) на картофеле;
- резкого снижения продуктивности пшеницы в США в 1917 г. из-за поражения стеблевой ржавчиной (Puccinia graminis tritici) и в 1950 г. из-за эпифитотии стеблевой ржавчины расы 15В;
- голода в Бенгалии в 1943 г. из-за эпифитотии гельминтоспориоза (Cochliobolus myabeanus) на рисе;
- гибели сортов овса, ведущих свое происхождение от сорта Виктория, из-за поражения Cochliobolus victoriae, и значительного снижения урожайности всех гибридов кукурузы с техасским типом цитоплазматической мужской стерильности (Т-цмс) в южных штатах США из-за поражения южным гельминтоспориозом (Helminthosporium turcicum, или Cochliobolus heterotrophus) в 1971 г.;
- внезапного распространения биотипа 1 бурой цикады (Niloparvata lugens) по сравнению с биотипом 2 после посева в 1974-1976 гг. на огромных площадях небольшого числа карликовых сортов риса на Филиппинах и в Индонезии.
Таким образом, опасения, вызванные возможностями уменьшения генетической изменчивости, полностью оправдываются, но их нельзя относить за счет только высокоурожайных сортов и тем более за счет технологии возделывания, потребностей рынка сбыта и финансовых доходов.
Нужно заметить, что всякое заселение и освоение новых территорий чужестранцами приводило к уменьшению числа и даже к уничтожению отдельных видов растений (трав, лесных пород и т.д.) и животных (птиц, насекомых и др.), но в то же время обусловливало распространение новых видов и разновидностей, необходимых для жизни людей и домашних животных, которые никогда прежде в природе так не распространялись. Примером может служить завоз в послеколумбовый период из Нового Света в Старый кукурузы, картофеля, томатов и табака или же из Старого Света в Новый пшеницы и других культур.
Высокоурожайные сорта и гибриды, созданные за последние годы в различных институтах стран мира, по ряду признаков не отличаются генетически ни друг от друга, ни от старых экстенсивных или местных сортов. Что касается устойчивости к патогенным организмам, то и здесь ни один местный сорт не обнаруживает каких-либо преимуществ. При изучении коллекции выращиваемых в Югославии сортов пшеницы не найдено местных сортов или популяций, устойчивых к ржавчине, за исключением двух, которые в действительности оказались американскими сортами Tatcher и Regent, завезенными в послевоенное время. Между местными сортами и патогенными организмами уже достигнуто равновесие на уровне средней устойчивости, вернее толерантности, что находит свое отражение в стабильной урожайности.
Следовательно, опасность возникновения эпифитотий существует не из-за недостатка различных сортов, а из-за желания фермера выращивать наиболее урожайный сорт, для чего в определенном районе и создают один или два сорта. Сколько лет уходит на создание по возможности большего числа генетически дивергентных сортов одной культуры, способных успешно противостоять основным лимитирующим факторам среды, столько же лет в одном районе, на одной ферме из-за меняющихся агроэкологических условий нужно выращивать ради устойчивой урожайности несколько генетически различных сортов.
Сохранение и использование наследственного материала
В настоящее время зародышевую плазму поддерживают в живом виде, и это необходимо по целому ряду причин. По мере развития науки и общества возникают новые потребности и новые возможности, и сохранение наследственного материала становится актуальным, о чем раньше даже не подозревали.
Для большей части человечества пшеница является не только основным продуктом питания, но и источником белка, поэтому возникла мысль о том, что с помощью селекций в ней можно повысить содержание белка. Улучшение питательной ценности также влечет за собой увеличение содержания лизина, которое в современных сортах пшеницы не превышает 3% в пересчете на сырой белок. В США изучение коллекции, включающей около 15 тыс. образцов, показало, что лишь несколько образцов пшеницы из Аргентины и Индии содержат около 4% лизина. Разумеется, эти образцы нужно сохранять и использовать в селекции как источник соответствующих генов.
До тех пор пока не была открыта мужская стерильность, которую можно использовать для получения гетерозисных гибридов растений, удаление тычиночных нитей с пыльниками (сорго, пшеница и т.д.) было невыгодным экономически, а найденные растения с мужской стерильностью выбрасывали за ненадобностью, не сохраняя их. Необходимость в генах-восстановителях фертильности и в использовании явления гетерозиса не возникала, поэтому никто не подозревал о различиях между генотипами по этому признаку и не искал их.
Что значит утрата селекционного материала, можно показать на одном из последних примеров. Несколько лет тому назад в Индию было послано из Югославии 10 новых высокоурожайных линий пшеницы. Там они прошли проверку на устойчивость к вирусным болезням, причем из всей испытанной коллекции только линия NS-879 оказалась устойчивой к возбудителю карликовой желтухи (YBDV). Когда из Индии об этом сообщили в Югославию и попросили прислать новые семена, это оказалось невозможным, так как в югославской коллекции этой линии уже не было.
Такие примеры существуют по каждой культуре, и можно суверенностью сказать, что сегодня безвозвратно теряется немало ценного селекционного материала, который может потребоваться завтра. Селекционерам об этом известно уже довольно давно, и многие страны еще в начале XX века, а в особенности после первой мировой войны обеспечили сохранность коллекций семян самых важных сельскохозяйственных культур. После второй мировой войны по мере интенсификации сельского хозяйства стали быстро исчезать местные популяции, стародавние и улучшенные сорта, а также вновь приобретенный и инорайонный селекционный материал. Поэтому в последние годы предпринимаются организационные меры, особенно со стороны ФАО, по поддержанию существующей генетической изменчивости культурных растений и их диких сородичей, для чего создаются так называемые генные банки.
К важнейшим источникам генетической изменчивости, которые необходимо сохранять, относятся следующие:
- сорта, которые возделывались и возделываются в производственных условиях;
- элитный селекционный материал, не прошедший официального испытания и районирования, но выделившийся по ряду признаков и находящий применение в программах гибридизации;
- селекционный материал, отличающийся специфическими признаками (устойчивость к болезням, высокое содержание белка и т.д.);
- местные популяции, в особенности те, генетическая изменчивость которых не использована при выведении селекционных сортов;
- специальный генетический материал (мутанты, генетические тестеры, источники цитоплазматической стерильности и т.д.);
- полиплоиды и анэуплоиды;
- синтетические гибриды;
- дикие сородичи.
Для сохранения такого материала необходимы хорошо организованные и оснащенные соответствующим оборудованием учреждения, позволяющие хранить семена многие десятки лет и освобождающие селекционеров от проведения частых пересевов. Система документации и информации должна быть организована таким образом, чтобы все заинтересованные лица могли быстро получать и использовать необходимые данные и материал.
На земном шаре существует несколько учреждений такого рода, где находятся на хранении коллекции семян и вегетативных частей растений. Наибольшее количество образцов имеют:
Всесоюзный институт растениеводства им Н. И. Вавилова, Ленинград, СССР;
Сельскохозяйственная исследовательская служба (ARS) министерства сельского хозяйства США, Белтсвилл, США;
Международный институт риса (IRRI), Лос Банос, Филиппины;
Международный институт сельскохозяйственных культур для полузасушливых тропиков (ICRISAT), Хайдерабад, Индия;
Международный центр по кукурузе и пшенице (CIMMYT), Эль Батан, Сьюдад Мехико, Мексика;
Голландско-немецкий генный банк по картофелю, Брауншвейг, ФРГ;
Международный центр по картофелю (IPC), Лима, Перу;
Международный институт сельского хозяйства тропиков (IITA), Ибадан, Нигерия;
Северный генный банк, Лунд, Швеция;
Азиатский центр по изучению и разработке овощных культур (AVRDC), Тайвань;
Эгейский региональный сельскохозяйственный институт (ARARI), Измир, Турция.
Кроме того, богатые коллекции хранятся в следующих национальных центрах:
Королевский ботанический сад, Лондон, Англия;
Институт селекции растений, Кембридж, Англия;
Национальный институт агрономических исследований (INRA), Версаль, Франция;
Центральный институт генетики и исследования культурных растений, Гатерслебен, ГДР;
Институт селекции сельскохозяйственных растений, Вагенинген, Голландия;
Национальный институт сельскохозяйственных наук, университет Киото, Япония.