Признак, генотип, фенотип
В предыдущих главах вкратце рассмотрены основные задачи генетики и фундаментальной селекции растений и различия между растениями-самоопылителями и перекрестниками. Показано, что ген - это основная единица наследственности, определяющая границы и направление развития определенного процесса и, в конце концов, определенного признака. Однако селекция ведется не на ген или гены, а на определенный признак, на фенотип. Поскольку для селекции живых организмов важнейшим является то, что передается по наследству, необходимо выявить связи между телом и признаком, между генотипом и фенотипом, а также между генотипом и факторами окружающей среды.
Признак
В генетике, а тем более в селекции организмов понятие признак, или особенность, употребляют для того, чтобы показать объективные различия между особями, вернее между сортами. Так, отличительными являются признаки окраски цветка (красная или белая), высоты стебля (высокий или низкий), устойчивости к болезням (устойчив или неустойчив), урожайности (высокоурожаен или низкоурожаен) и т.д.
Таким образом, проявление признака, или особенности, есть определенная характерная черта фенотипа. Любая особь, любой генотип обладают огромным числом признаков, границы которых, однако, не всегда легко установить. Поэтому генетик воспринимает признаки несколько иначе, чем селекционер, селекционер - иначе, чем биохимик, и т.д.
В основе любого признака лежит отдельный ген или комплекс генов, которые определяют границы развития самого признака. В этом и заключена генетическая сторона признака, т.е. то, что определяет генотип. Кроме того, формирование каждого признака - это закономерный результат действия факторов внешней среды, которые всегда варьируют и модифицируют сам признак. Йогансен установил, что как фенотип представляет собой конечный продукт проявления общего действия генотипа и среды обитания, так и любой признак обусловлен влиянием генетических факторов и факторов окружающей среды. Доля наследуемого, или генетического компонента, как и доля ненаследуемого, или экологического компонента, обусловленного средой, различна для любого определяемого признака, и установить ее всегда нелегко. Для селекции в первую очередь важное значение имеет генетический компонент признака, т.е. тот, который передается по наследству потомству. Это особенно характерно для количественных признаков, имеющих в большей или меньшей мере скрытую изменчивость, которая обнаруживается вследствие влияния условий среды обитания и не наследуется.
Например, на человека постоянно производит впечатление крупность колосьев, початков или плодов растений, находящихся в крайних рядках делянки, и он не может удержаться, чтобы не отобрать их. На следующий год потомство этих «лучших» растений, как правило, уступает потомству растений, отобранных из срединных рядков делянки. Следовательно, произошел отбор модификаций, появившихся под влиянием более благоприятных условий вегетации, повышенной фотосинтетической активности и т.д., что относится к ненаследственной изменчивости, не передающейся потомству. Степень контроля генетического компонента изменчивости в ходе отбора на какой-то признак зависит от количества генов, определяющих данный признак, их эффекта и от силы влияния факторов среды.
Признаки, обусловленные главными генами, или генами с мощным эффектом, т.е. окраска цветков и плодов, форма цветков, листьев, плодов, зерен и т.д., обычно легко различить на глаз, и в результате потомство чаще всего характеризуется отобранным признаком. Однако, если главный ген обладает доминантным эффектом, одинаковые фенотипы не должны иметь тождественный генотип. Например, отобраны два растения с красной окраской цветков, т.е. одного и того же фенотипа. В потомстве одного растения все особи имеют красную окраску, в потомстве другого получены растения и с красной, и с белой окраской цветков. Это значит, что первое растение было гомозиготным (СС), а второе - гетерозиготным по красной окраске (Сс).
Если речь идет о большом количестве генов, обусловливающих проявление одного признака, то одинаковые фенотипы могут содержать различные гены. Например, у одних сортов тыквы признак округлой формы плода обусловлен действием генов ААbb, у других - действием генов ааВВ. Существует множество других типов взаимодействия генов в детерминировании разных признаков, о чем говорилось в главе об источниках изменчивости.
Как несколько генов могут обусловливать развитие одного признака, так и один единственный ген может оказывать влияние на несколько признаков. В последнем случае имеются в виду гены с плейотропным, или многосторонним действием, например ген фиолетовой окраски стебля и чашелистиков у Primula sinensis и других растений.
Кратчайший путь между первичным действием гена и его конечным выражением в фенотипе несложно пронаблюдать на взаимоотношении ген - признак. Серповидная анемия человека - следствие замены одного основания в кодоне (GAA на GUA); вместо глутаминовой кислоты в B-цепь в положение 6 включается валин, и это обусловливает изменение гемоглобина. В данном случае налицо прямая связь между геном и признаком. Однако у огромного числа признаков, особенно тех, с которыми работает селекционер, от первичного действия гена до его выражения в признаке генотипа проходит весьма длительный процесс, и приводит он к взаимодействию с другими генами, часть которых оказывает влияние в одной, а часть - в другой фазе развития признака и организма как единого целого. Если к этой совокупности эффекта гена добавляется эффект факторов среды, модифицирующих действие гена, то возникновение связи между геном и признаком заметить всегда трудно. Во всяком случае, это нелегко в отношении таких количественных признаков, как содержание белка, масса плода, урожай зерна и т.д.
Собственно говоря, с точки зрения количественных показателей понятие «признак» представляет в большей мере агрономическую или селекционную, нежели генетическую, категорию. Кроме того, к признаку, имеющему хозяйственно важное значение, все чаще и чаще приходится подходить комплексно. Действительно, урожай можно рассматривать сейчас не как отдельно взятый признак, а как совокупность признаков. Урожай зерна пшеницы и других зерновых культур состоит из таких структурных элементов, как число растений (колосьев) на 1 м2, число зерен в колосе и абсолютная масса зерна. Каждый из этих элементов можно рассматривать как отдельно взятый признак, но вместе они дают конечный продукт - урожай зерна. Матер и Джинкс называют эти элементы структуры урожая подпризнаками, а сам урожай зерна - сверхпризнаком.
Эффект гена и факторов окружающей среды обусловливает непрерывную изменчивость признака. Поэтому, если нельзя отличить генетическую изменчивость от негенетической, следует всегда проводить только испытание по потомству. Кроме того, необходимы определенные эксперименты, позволяющие установить в суммарной фенотипической изменчивости взаимодействие между генами и взаимодействие генов с окружающей средой.
Фенотип и компоненты фенотипической изменчивости
Количественный признак фенотипа устанавливают путем измерения. Следовательно, полученное его значение представляет собой фенотипическую ценность анализируемых особей, т.е. это совокупная ценность, состоящая из генотипической ценности особи и отклонений, вызванных действием факторов окружающей среды. Выразить ее можно следующим образом:
F (фенотип) = G (генотип) + Е (влияние экологических факторов).
Отдельные особи объективно различаются по фенотипической ценности. Эти различия обусловлены наличием генетических различий между данными особями, влиянием факторов среды и взаимодействием между генотипом и факторами среды. Таким образом, фенотипическая ценность изменчива и состоит из компонентов, которые можно установить путем анализа вариансы. Так, фенотипическая изменчивость включает генотипическую изменчивость, изменчивость, вызванную влиянием факторов окружающей среды (экологическую изменчивость), и их взаимодействие:
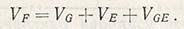
Источник генотипической изменчивости (VG) заключен в генетической конституции самого количественного признака. Если гены обнаруживают аддитивный эффект, то при замене одного из них генотипическая ценность такого признака будет или увеличиваться, или уменьшаться. Например, если ценность A1A1 равна 6 см, A1A2 - 7 см и A2A2 - 8 см, то это означает, что присутствие гена A2 является причиной изменения на 1 см. Отдельные гены могут даже обладать доминантностью, но присутствие одного аллеля может вызвать увеличение значения генотипической ценности. В таком случае генотип A1A2 будет иметь ценность, равную не 7, а 8 см. Возможно также взаимодействие между различными аллелями, или так называемый эпистаз. Допустим, что Аа обладает аддитивным эффектом, действуя вместе с ВВ, но с bb обнаруживает доминантность. Это указывает на наличие генотипической изменчивости, определяемой компонентами, которые можно выразить следующим образом:
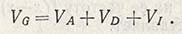
Следовательно, генотипическая варианса включает вариансы с аддитивным и доминантным действием генов и с взаимодействием между ними, т.е. фенотипическая изменчивость состоит из:
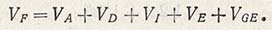
Величины значений отдельных компонентов вариансы оценивают в экспериментах. Если все особи имеют один и тот же генотип, то изменчивость, устанавливаемую в эксперименте, можно отнести за счет влияния факторов среды. Сходные генотипы могут быть самоопылителями или инбредной линией, которая практически гомозиготна. Поколение F1 полученное от скрещивания двух самоопыленных линий, генетически однородно, хотя и гетерозиготно. Поэтому вариансы родительских форм и F1 могут быть использованы в качестве мерила экологической вариансы (VE).
Для разложения генотипической вариансы на отдельные компоненты используют вариансу F2 и поколения возвратных скрещиваний. Одним из первых этот способ разработал Матер. Поскольку в F2 происходит расщепление по признакам, варианса этого поколения включает вариансу каждого генотипа, а также вариансу, возникшую под действием факторов среды. Например, при наличии только одной пары генов (A1 и A2) в F2 появляются три генотипа в соотношении:
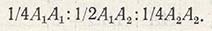

Каждый из этих генотипов обладает генотипической ценностью, которая представляет собой некоторое отклонение от средней для всего родительского поколения:
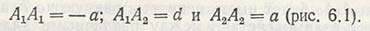
Подставив эти значения в вышеприведенное соотношение, получим следующую среднюю ценность F2:
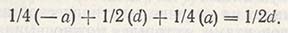
Варианса любого генотипа равна квадрату отклонения от средней ценности, помноженной на ее частоту f(x-x)2, следовательно, суммарная варианса F2 составляет:
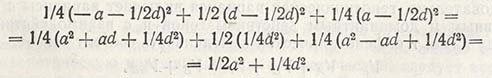
Если а2 заменить буквой A, a d - буквой D и добавить компонент вариансы, полученной под влиянием среды (Е), то окажется, что итоговая варианса F2 равна:
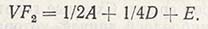
Эти компоненты фактически представляют собой вариансы аддитивности (VA), доминантности (VD) и влияния среды (VE). Таким образом, компоненты вариансы родительского поколения (Р1, Р2) и поколений, в которых чаще всего проводят оценки (F1 и F2), а также вариансы возвратного скрещивания с первым (B1) и вторым (В2) родителями можно выразить следующим образом:
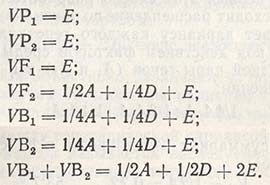
Способ расчета компонент вариансы можно рассмотреть на примере наследования числа колосков в колосе при скрещивании мексиканского сорта пшеницы Siete Cerros и советского сорта Безостая 1 (табл. 6.1). Прежде всего вычисляют вариансу действия факторов среды, которая включает вариансы поколений родителей и F1:
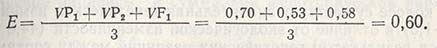
Если значение Е (0,60) вычесть из значения суммарной вариансы F2 (1,34) и значение 2Е (2x0,60) - из средней вариансы возвратных скрещиваний (2x0,98), остаются только вариансы аддитивности и доминантности:
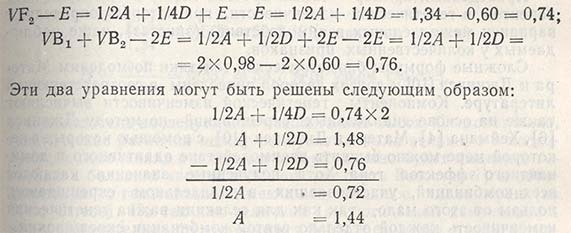
Подставив полученное значение в верхнюю часть уравнения, можно вычислить значение D:

Таким образом, суммарная изменчивость в F2 состоит из следующих компонентов:
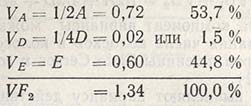
Данный анализ показывает, что в F2 по признаку числа колосков в колосе существует значительная генетическая изменчивость (53,7 + 1,5) в отличие от экологической изменчивости (44,8%); это является результатом генетических различий между сортами Siete Cerros и Безостая 1 (табл. 6.1). Более того, наибольшая доля генетической изменчивости приходится на аддитивное действие генов (53,7%) и очень малая - на доминантное (1,5%). Приведенный пример - самый простой способ вычисления компонентов в генетической изменчивости, при котором определяют вариансу межаллельных взаимодействий (эпистаз), чаще наблюдаемых у количественных признаков.
Сложные формулы биометрической генетики по моделям Матера и Джинкса и других авторов приведены в соответствующей литературе. Компоненты генетической изменчивости вычисляют также на основе диаллельных скрещиваний по методу Джинкса, Хеймана, Матера и Джинкса, с помощью которых в некоторой мере можно выявить взаимодействие аддитивного и доминантного эффектов гена. Хотя полученные значения касаются всех комбинаций, участвовавших в диаллельном скрещивании, пользы от этого мало, так как для селекции важна генетическая изменчивость каждой отдельно взятой комбинации скрещивания.
Наследуемость
Отбор проводится на основе фенотипической ценности, поэтому важно знать, с какой вероятностью отобранные фенотипы дадут тождественное потомство. Если значение генетической вариансы по определенному признаку велико, а значение экологической вариансы мало, можно ожидать, что потомство в немалой мере будет таким же, как и отобранные фенотипы. И наоборот, если генетическая варианса мала, а экологическая велика, потомство по ценности может значительно отличаться от отобранных фенотипов.
Сходство между родителями и их потомством в огромной степени зависит от компонентов генетической вариансы (VA + VD). Если речь идет об аддитивном компоненте генетической вариансы (VA), то здесь фенотипы родителей представляют собой надежные индикаторные признаки своих генотипов и, следовательно, дадут сходное потомство. При доминантном компоненте генетическая варианса (VD) будет давать отличное от родительских фенотипов потомство, и это зависит от характера межаллельного взаимодействия.
Соотношение между генотипической вариансой и суммарной фенотипической вариансой называется наследуемостью (Н, или h2) какого-то признака определенной популяции и обозначается:
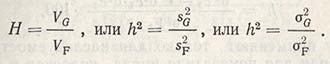
Это наследуемость в широком смысле. Наследуемость в узком смысле - соотношение только между аддитивным компонентом генотипической вариансы и суммарной фенотипической вариансой:
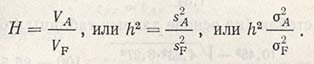
Наследуемость по признаку числа колосков в колосе в проанализированном примере составляет:
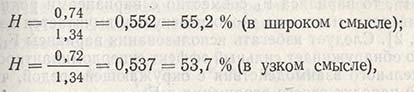
т.е. показатель наследуемости относительно высок. Следовательно, генетические различия между родителями были велики, и в последующих поколениях с помощью отбора можно будет выделить генотипы с большим числом колосков (из сорта Безостая 1) и объединять их с генотипами, имеющими большое число зерен (из Siete Cerros). Но так как величина экологической изменчивости весьма значительна, то этого может быть достаточно, чтобы истинная ценность генотипов оказалась скрытой и произошел отбор модификаций, которые в следующем поколении не дадут растений с большим числом колосков.
Как уже было сказано, для успешного проведения отбора наибольшее значение имеет аддитивный компонент генетической изменчивости, названный поэтому селекционной ценностью. Фальконер считает, что наследуемость выражается в пригодности фенотипической ценности служить ориентиром селекционной ценности или же она отражает степень совпадения между фенотипической и селекционной ценностью.
Другие методы расчета наследуемости
Полнее всего, как показано выше, наследуемость после гибридизации рассчитывают по формулам Матера. Наследуемость в широком смысле можно также рассчитать только по F2, если допустить, что окружающая среда в одинаковой мере влияет и на поколение родителей, и на популяцию F2. Разница между средней ценностью варианс поколения родителей и F2 дает генотипическую вариансу. Наследуемость вычисляют по формуле:
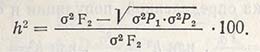
Эту формулу применяют только для наследуемости в широком смысле, которая для показателя числа колосков в колосе составляет:

Если вместе с F2 и поколениями родителей в те же годы выращивают F1, то варианса F1 совместно с вариансами родительских поколений принимается за экологическую и вычитается из вариансы F2. Следует избегать использования вариансы F1, так как она часто обнаруживает сильный эффект супердоминантности и последовательного взаимодействия с окружающей средой, что почти всегда не находит своего отражения в F2.
Наследуемость можно рассчитывать и как регрессию селекционной ценности от фенотипической ценности:

Наследуемость также можно рассчитать и через коэффициент корреляции между селекционной и фенотипической ценностью:
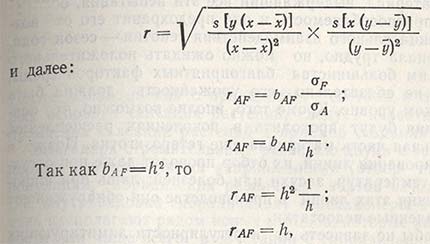
а это означает, что коэффициент корреляции между селекционной ценностью (А) и фенотипической ценностью (F) равен наследуемости. Следовательно:
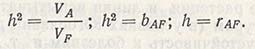
Выведение формул показано в учебниках Фальконера, Матера и Джинкса и других авторов.
Учитывая, что наследуемость отдельных признаков имеет огромное значение для генетической ценности отбора, о ней еще будет сказано в главе о методах селекции.
Взаимодействие генотип-среда в процессе селекции
Выше была рассмотрена роль компонентов генетической изменчивости и соотношения генетической и экологической изменчивости в выраженности одного единственного признака. Однако как между отдельными признаками и факторами окружающей среды, так и между генотипом как единым целым (особенно в отношении урожая) и факторами окружающей среды может произойти взаимодействие (VGE), что следует учитывать в ходе процесса селекции.
Создание новых сортов растений - обычно процесс длительный, и селекционный материал подвергается действию факторов окружающей среды на протяжении большого числа лет. На создание и передачу в производство нового сорта однолетних растений в среднем уходит около 10 лет, для многолетних растений - значительно больше.
Начиная с F2, проводят отбор фенотипов, у которых, как ожидается, произойдет рекомбинация генов с проявлением положительных агрономических признаков. Вследствие сильного ежегодного варьирования условий среды один год может быть благоприятным для испытаний на засухоустойчивость, второй - для оценки на устойчивость к низким температурам, третий - для испытаний на устойчивость к болезням и т.д. После 5-6 лет отбора можно ожидать, что материал, выдержавший все эти испытания, обнаружит широкую приспособляемость и она предохранит его от возникновения отрицательного взаимодействия генотип - сезон года. У такого материала трудно, но можно ожидать положительного использования им большинства благоприятных факторов среды, причем совсем не обязательно его урожайность должна быть на самом высоком уровне. Кроме того, вполне возможно, что повторные испытания будут проходить в поколениях расщепления, когда значительная часть материала еще гетерозиготна. Позже, в процессе формирования линий, их отбор проводят даже при отсутствии низких температур, засухи или болезней; лишь при широком использовании этих линий в производстве они обнаруживают ранее неустановленные недостатки.
Поэтому, чтобы не зависеть от нерегулярности лимитирующих факторов окружающей среды, в процессе отбора принято создавать искусственные условия (используя теплицы, фитотроны, лаборатории) и материал в поколениях расщепления, а также первоначально отобранные растения и линии испытывать на устойчивость к низким температурам (в условиях Югославии до -15°С), на засухоустойчивость, устойчивость к болезням и т.д. Для более полной проверки влияния климата и патогенных организмов значительное число селекционных учреждений выращивает материал в расщепляющихся поколениях и проводит отбор по крайней мере в двух различных географических районах, что в немалой степени может заменить сезоны года. Все эти испытания снижают возможность возникновения риска от неблагоприятного взаимодействия генотип - окружающая среда.
О взаимодействии генотип - среда будет более подробно рассказано в главе о приспособляемости и стабильности сортов.