Активная, или истинная, генетическая устойчивость
Генетическая устойчивость обеспечивает истинную, или активную, устойчивость растений-хозяев к патогенам. Устойчивое растение в действительности сверхвосприимчиво; используя продукты своего обмена веществ (полифенолы, токсины), оно сдерживает инвазию патогеном или мешает его успешному развитию. Это означает, что такое растение содержит гены устойчивости, продукты действия которых в состоянии воспрепятствовать прониканию патогена и не дать ему развиваться. В то же время это означает, что патоген не обладает генами вирулентности, которые дали бы ему возможность совершить инвазию растения-хозяина, и соответственно обладает генами авирулентности. Наоборот, если патоген имеет гены вирулентности, а у растения отсутствуют гены устойчивости, развивается болезнь.
Существование в природе некоторого специфического механизма взаимоотношений патогена и хозяина, о котором здесь идет речь, подтверждено исследованиями Стакмана и др. в США. Уже в начале XX века они приступили к изучению популяции ржавчины и установили, что различные сорта пшеницы по-разному реагируют на два изолята Puccinia graminis tritici.
В соответствии с проведенными исследованиями сорта растений обладают различными генетическими структурами в отношении устойчивости к патогену, а популяции патогенов обладают различными генетическими структурами в отношении вирулентности или способности осуществлять инфекцию. Полученные данные открыли дорогу для новых исследований, позволивших выявить огромную генетическую изменчивость или, другими словами, многочисленные физиологические расы патогенов внутри популяций отдельных видов патогенов. При испытаниях культур патогенов различной степени вирулентности на сортах-хозяевах у растений были обнаружены многочисленные гены устойчивости и восприимчивости. Чтобы результаты подобных исследований можно было сравнивать, введена международная система идентификации генов устойчивости у хозяина и физиологических рас у патогена.
К анализу популяций патогенов подошли путем тестирования различных сортов растений. Такой сортимент, т.е. сорта-дифференциаторы, с помощью которых сравнивают результаты тестов, установлены на международном уровне. Очевидно, чем больше сорта различаются по степени устойчивости, которую можно идентифицировать, тем большим является число рас патогена и соответствующих генов устойчивости хозяина. Например, тест-сортимент для листовой ржавчины пшеницы включает 12 сортов, для мучнистой росы пшеницы - 14, для фитофтороза картофеля - 9 сортов-дифференциаторов и т.д. Современные генетические знания о взаимосвязях паразита и хозяина настолько велики, что сегодня для анализа популяций патогена вместо сортов-дифференциаторов все чаще используют изогенные линии, обладающие отдельными генами устойчивости к соответствующим видам патогенов и имеющие сходную генетическую основу по остальным признакам. Так, для идентификации устойчивости к листовой ржавчине пшеницы используют изогенные линии сорта Thatcher с различными генами Lr.
Идентификация рас основывается на типах реакций или типах инфекции. У зерновых оценку устойчивости к ржавчине проводят на основе типа инфекции и процента интенсивности поражения (severity) (табл. 14.1).

Типы инфекции от 0 до 4 в основном применяют при оценке всходов в теплице. Добавление рядом с цифрой знака (+) или (-) дает возможность подробнее описать тип реакции. В полевых условиях типы реакции взрослых растений чаще всего обозначают буквами R (устойчивость), MR (умеренная устойчивость), MS (умеренная восприимчивость) и S (восприимчивость).
Очень важное значение при проведении такой работы в полевых условиях имеет установление интенсивности поражения, оцениваемой от 0 до 100%. Например, оценка 10R, 25MR, 60S означает, что 10, 25 или 60% листовой поверхности характеризуется устойчивостью, умеренной устойчивостью и восприимчивостью (рис. 14.1).

Показатели типов реакций и интенсивности поражения используют для расчета коэффициентов, на основе которых устанавливают степень устойчивости растений-хозяев.
Оценку устойчивости, точнее восприимчивости к некоторым другим патогенным организмам, у которых труднее различить тип инфекции, осуществляют главным образом путем установления степени поражения с использованием шкалы баллов от 0 до 5 или от 0 до 9 (0 - отсутствие заражения, 1 - устойчивость, 5 - относительная восприимчивость, 9 - высокая восприимчивость растений).
На основе типов поражения сортов-дифференциаторов и с помощью международных ключей к настоящему времени идентифицировано огромное количество физиологических рас патогенов, в том числе:

Число физиологических рас, которое можно идентифицировать в популяции какого-либо вида патогена, зависит от числа сортов-дифференциаторов растений-хозяев и числа уровней взаимодействия патогена и хозяина (R, MR, MS, S и т.д.). Согласно Ван дер Планку, его можно рассчитать по формуле:
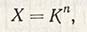
где X - число физиологических рас; К - число уровней взаимодействия между патогеном и хозяином; n - число сортов-дифференциаторов с различными генами устойчивости.
Точно так же идентифицируют гены устойчивости хозяина. Например Sr1...Sr32 - гены устойчивости к стеблевой ржавчине пшеницы, гены Lr1...Lr26 - к листовой ржавчине пшеницы, Pg1...Pg13 - к стеблевой ржавчине овса, R1...R4 - гены устойчивости к фитофторозу картофеля и т.д. Очень обстоятельные сведения об этом приведены в работе Макинтоша.
Генетическое объяснение устойчивости к патогенам впервые дал Флор, выдвинувший теорию ген-на-ген. Так, если растение-хозяин имеет ген устойчивости, то определенная раса патогена не в состоянии вызвать инфекцию. В данном случае сорт устойчив к этой расе патогена, а сам патоген авирулентен. Если же раса патогена в состоянии вызвать инфекцию у определенного сорта, тогда патоген имеет ген вирулентности, а растение-хозяин не имеет гена устойчивости и является неустойчивым.
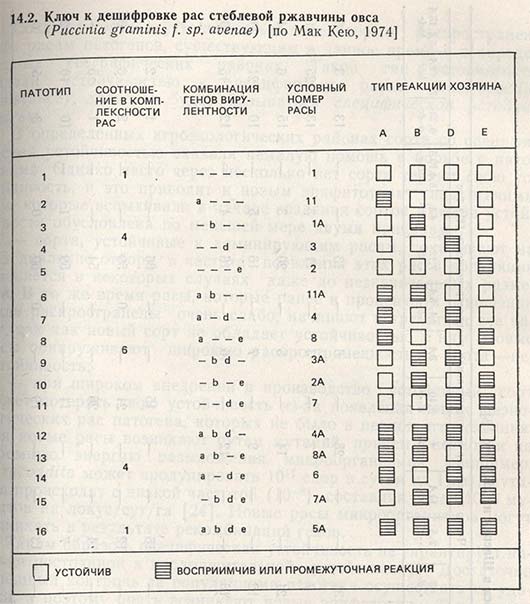
Простым и наглядным примером доказательства теории ген-на-ген служит идентификация рас стеблевой ржавчины овса (табл. 14.2). У четырех сортов овса: Richland (носитель гена A), Rodney (В), White Tortor (D) и Sevenothree (Е) идентифицировано 16 рас или патотипов Puccinia graminis avenae. Полученные данные показывают, что если один сорт может быть устойчив к одним и восприимчив к другим расам, следовательно, существуют и расы, не имеющие генов вирулентности или имеющие только один ген, но тем не менее различимые по двум или трем различным генам вирулентности или по всем четырем генам вирулентности.
Расы патогена по-разному распределены в пространстве и во времени, о чем весьма наглядно свидетельствуют данные таблицы 14.3. Некоторые расы P. graminis avenae (7, 12, 6, 13) по сравнению с другими (ЗА, 4А) представлены очень широко, но процент их распространенности может различаться по годам. Что касается распределения рас в пространстве, то большинство их обнаружено в Швеции и США, причем одни встречаются только в Швеции (3, 4, 3A), а другие - только в США (2, 5, 8, 10).
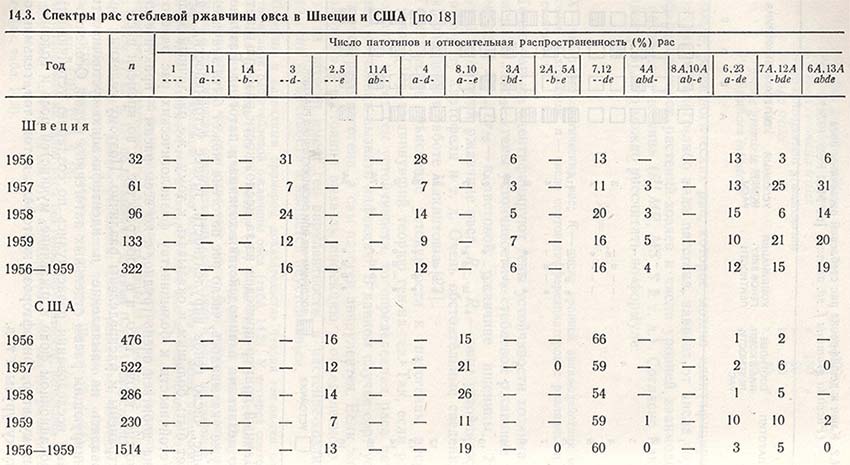
Не установлено ни одной расы без генов вирулентности, небольшое их число (3 и 2,5) имеет один ген (d или е), а большинство - два или три различных гена вирулентности; только две расы (6A, 1ЗA) имеют четыре гена вирулентности (abde) (см. табл. 14.3).
Данный пример приведен из-за несложности разбора генетического механизма взаимодействия хозяина и патогена. Разумеется, как уже указывалось, число рас патогена может быть значительно больше, число генов вирулентности, точнее устойчивости, также может быть большим и оказывать к тому же различный эффект.
Устойчивости к большинству физиологических рас одного патогена добиться очень трудно, а ко всем расам - практически невозможно; кроме того, и в пространстве, и во времени расы распространены и распределены различно. Поэтому селекция ориентировалась на выявление и идентификацию устойчивости к Доминирующим расам отдельных патогенных видов. Особенно усиленные исследования развернулись по созданию устойчивости на провокационном фоне к ржавчине, мучнистой росе и головне зерновых культур, фитофторозу картофеля и томатов, гельминтоспориозу кукурузы и т.д.
Созданы сорта с устойчивостью к некоторым распространенным расам патогенов, существующим в данное время и в определенных географических районах. Такой тип устойчивости назван устойчивостью к специфичным расам (race-specific resistance), далее он будет называться специфической устойчивостью.
В определенных агроэкологических районах сорта со специфической устойчивостью оказали немалую помощь в борьбе с патогенами. Однако часто через несколько лет сорта теряют свою устойчивость, и это приводит к новым эпифитотиям, аналогичным тем, которые вспыхивали в начале создания сортов. Потеря устойчивости обусловлена по меньшей мере двумя причинами:
- сорта, устойчивые к доминирующим расам, оказывают на них давление отбора, и частота появления этих рас в популяции снижается в некоторых случаях даже до незначительных размеров. В то же время расы, которые ранее в процентном отношении были распространены очень слабо, начинают встречаться все чаще, так как новый сорт не обладает устойчивостью к ним. Новые расы обнаруживают широкую распространенность, а сорта - неустойчивость;
- при широком внедрении в производство устойчивый сорт может потерять свою устойчивость из-за появления новых физиологических рас патогена, которых не было в период его создания. Эти новые расы возникают путем мутаций, причем, несмотря на огромную энергию размножения микроорганизмов (например, P. recondita может продуцировать 10й спор в сутки на 1 га), мутации происходят с низкой частотой (10-8), составляя около 1000 мутантов на локус/сут/га. Новые расы микроорганизмов могут возникать в результате рекомбинаций генов.
Таким образом, специфическая устойчивость не гарантирует наличия постоянной и продолжительной устойчивости. Достаточно успешный контроль за популяциями патогена осуществить не удается, и поэтому опять возникают новые эпифитотии. Так, после усиленной работы по идентификации физиологических рас и созданию сортов пшеницы, устойчивых к доминирующим расам P. graminis tritici, в США и Канаде в 1935 г. появилась новая раса 56, в 1953 г. - раса 15В, а затем в 1954 г. разразилась огромная эпифитотия, снизившая урожаи в отдельных районах приблизительно на 50%.
В то же время замечено, что некоторые сорта, не имеющие генов устойчивости к доминирующим расам, в течение длительного времени обладают относительно умеренной, средней степенью устойчивости.
Весь накопленный опыт побудил ученых к дальнейшим размышлениям над собранными фактами и к новым исследованиям с целью выяснения, с чем же в действительности должна вестись работа. В результате Ван дер Планк выдвинул новую теорию о вертикальной и горизонтальной устойчивости.