Наследование качественных признаков
Качественные (альтернативные) признаки - это признаки, развитие которых обусловлено действием генов с сильным эффектом, или так называемых главных генов. К подобным признакам относятся окраска и форма цветков, плодов, листьев и других органов, в ряде случаев устойчивость к патогенным организмам и т.д.
Идеальные сорта, т.е. обладающие комбинациями генов и признаков, которые могли бы долгие годы удовлетворять соответствующим требованиям, не существуют, и поэтому постоянно возникает потребность в создании новых комбинаций генов и новых сортов, по своим признакам отличающихся от всех предыдущих сортов.
Для комбинирования генов сорта-родители должны различаться по меньшей мере двумя парами генных локусов, точнее двумя признаками. Желательно, например, при выведении сорта ячменя добиваться того, чтобы он был двурядным и устойчивым к ржавчине. В Югославии есть двурядный сорт ячменя, но неустойчивый к ржавчине, и сорт, устойчивый к ржавчине, но шестирядный.
Из предыдущих исследований известно, что ген, ответственный за признак двурядности, доминантен (D) по отношению к гену шестирядности (d), а ген устойчивости к ржавчине доминантен (R) по отношению к гену неустойчивости (r). К скрещиванию приступают, прежде всего установив, какой сорт будет служить материнским и какой - отцовским (т.е. выбрав материнское и отцовское растение). Если в качестве материнского отобран двурядный, неустойчивый к ржавчине сорт, это означает, что потребуется время на удаление (кастрацию) пыльников, чтобы предотвратить самоопыление. У ячменя, имеющего три пыльника, их необходимо удалять пинцетом с каждого цветка в колосе еще до созревания; в условиях Югославии это происходит в мае, когда колос находится в пазухе верхнего листа (до выколашивания). Через 2-3 дня на такой кастрированный цветок, имеющий только пестик, наносят зрелую пыльцу, собранную из пыльников отцовского - шестирядного, устойчивого к ржавчине сорта. Таким образом, схема гибридизации и развития дальнейших поколений будет иметь следующий вид:
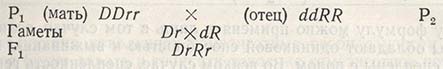
В F1 развиваются двурядные растения, устойчивые к ржавчине, но их родители различались по двум парам аллельных генов. Поколение F1 гетерозиготно, и эти растения не представляют сорта, так как в F2 произойдет расщепление. Следовательно, нужно получить растения F2, что у ячменя сделать просто: без кастрации или какого-либо вмешательства, поскольку каждое растение F1 самофертильно.
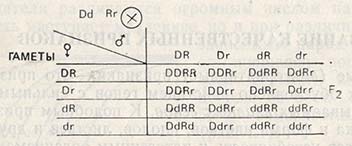
За счет рекомбинаций генов из 16 возможных возникнут 9/16 желательных комбинаций, т.е. двурядные, устойчивые к ржавчине ячмени. Однако все эти фенотипы имеют разные генотипы: 1/16 гомозиготна по обоим парам генов (DDRR), 2/16 гомозиготны по гену двурядности, но гетерозиготны по гену устойчивости (DDRr), 4/16 гетерозиготны по обоим генам (DdRr) и 2/16 гетерозиготны по гену двурядности и гомозиготны по гену устойчивости (DdRR).
Поскольку ген двурядности и ген устойчивости обнаруживают полную доминантность над своим рецессивным аллелем, гомозиготное состояние от гетерозиготного по фенотипу отличить нельзя, и поэтому в F2 нужно отобрать все растения, обладающие двурядностью и устойчивостью (D-R-). Гомозиготные по обоим желаемым признакам генотипы можно отобрать лишь в F3. Это будут потомства, у которых ни по признаку двурядности, ни по признаку устойчивости (DDRR) расщепления не произойдет. При наличии достаточного количества таких растений отпадает необходимость в отборе двурядных и устойчивых форм с других делянок, однако при этом нужно отобрать все растения с генотипом DDRR и высеять поколение F4.
Если в F4 не происходит расщепления ни по одному из признаков, то можно сказать, что получена двурядная, устойчивая к ржавчине линия ячменя (DDRR). В дальнейших поколениях эту линию размножают, и если она отвечает требованиям показателей урожайности и качества, новый сорт можно считать созданным.
В данном случае речь шла только о двух главных генах, а обнаруженная рекомбинация содержала оба доминантных парных гена, т.е. частота проявления этих фенотипов в F2 была значительно выше, чем у всех остальных фенотипов (9/16 от вместе взятых 16/16). Таким образом, выделение ценных рекомбинаций позволяет избежать формирования большой популяции растений в F2, достаточно сформировать популяцию всего лишь из нескольких сотен растений. Например, популяция из 300 растений содержала 9/16x300=168 растений двурядных и устойчивых к ржавчине (D-R-), из которых 18 растений имели генотип DDRR (1/16x300). Этого числа достаточно, чтобы в дальнейших поколениях выделить необходимое число линий с ценными рекомбинациями.
Приведенный пример предполагает простейший случай наследования признаков и рекомбинаций генов. На практике часто возникает потребность в рекомбинациях по трем и более признакам, например, когда ставят целью создать сорт ячменя двурядного (DD), устойчивого к ржавчине (RR), с прямым типом колоса (nn). Поскольку сорта с такими признаками нет, скрещивают два или больше сортов, обладающих указанными особенностями:
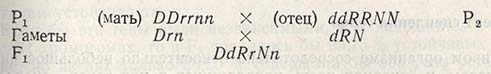
Поколение F1 содержит двурядные и устойчивые растения с нужным типом колоса. Полученную ценную рекомбинацию размножают до F2.
Растения F1 с генотипом DdRrNn дадут восемь типов гамет: DRN, DRn, DrN, Drn, dRN, dRn, drN и drn. От свободного оплодотворения этих гамет в F2 образуется 64 комбинации, из которых 9/64 будут содержать требуемые рекомбинации (D-R-nn). В ходе работы нужно отобрать двурядные устойчивые растения с прямым типом колоса. Признак прямого колоса рецессивен, и расщепления ни в одном из потомств по этому признаку не произойдет; что же. касается рядности колоса и устойчивости, то в одних потомствах. расщепление возможно, в других - нет. Поэтому необходим дальнейший отбор по этим двум признакам до тех пор, пока не будет достигнута гомозиготность.
Когда речь идет о трех признаках и поисках рекомбинаций генов, теоретически можно предположить, что для репрезентативности с частотой 9/64 в F2 необходимо получить большее число растений, чем для случая с двумя признаками. Если бы в F2 популяция насчитывала 300 растений, то частота появления ожидаемых рекомбинаций составила 9/64X300 = 42 растения, т.е. небольшое число. Однако иногда развиваются не все растения, а часть их просто не замечают при отборе, и в результате шансы на успех падают. Поэтому в F2 нужно было бы иметь около 1000 растений, что обеспечило бы получение 140 растений с ценными рекомбинациями генов (9/64Х1000). Среди такого числа растений можно наверняка найти достаточно особей для ведения дальнейшей работы.
Эффект взаимодействия генов
В предыдущих примерах рассмотрен случай с независимым действием генов, когда два и более генных локуса находятся в различных хромосомах и каждый независимо друг от друга влияет на развитие какого-то признака.
Между тем ряд генных локусов находится во взаимодействии, имеет различный эффект и даже приводит к иному соотношению расщепления в F2, а именно: 9:7, 9:3:4, 9:6:1, 3:13, 15:1 и т.д. Об этом уже шла речь в главе 3, где обсуждалось возникновение новой генетической изменчивости после гибридизации. В таких случаях предвидеть желаемые рекомбинации генов трудно или даже невозможно, особенно при отсутствии более ранних сведений из литературных источников. И тем не менее получить совершенно новые признаки все-таки можно.
Эффект сцепления генов
В живом организме сосредоточено относительно небольшое число хромосом, и основная часть находящихся в них генов сцеплена. При сцеплении генов, определяющих два положительных агрономических признака, данная родительская комбинация представляет ценность, но если в одном сорте сцеплен один положительный (и один отрицательный признак, необходимо провести рекомбинацию генов с другим родителем. В этом случае вследствие сцепленности генов (linkage) в потомстве после скрещивания появляется самое большое число родительских комбинаций генов, а рекомбинации могут произойти только в результате слияния кроссинговерных гамет, Число рекомбинаций зависит от отдаленности генных локусов друг от друга. Если речь идет о двух сцепленных и достаточно отдаленных генных локусах, то ожидаемые рекомбинации можно получить в относительно небольшой по численности популяции F2. Если же сцеплено три (и более) генных локуса, а их отдаленность невелика, то необходима очень большая по численности популяция, и в данном случае ожидаемые рекомбинации получить чрезвычайно трудно (табл. 4.4).
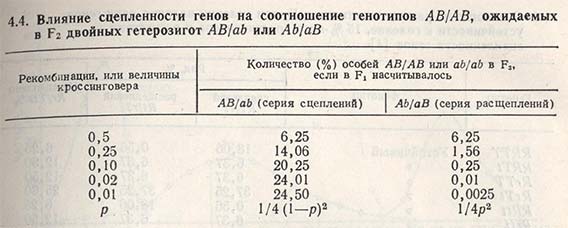
Меру влияния сцепленности генных локусов на ожидаемые рекомбинации генов и различия между рядами сцепления и расщепления в сравнении с несцепленными генными локусами можно показать на примере наследования пшеницей признака устойчивости к головне.
Установлено, что признак устойчивости к головне обусловлен двумя генами (Т и R), сцепленность которых составляет приблизительно 15% кроссинговера. Если оба гена устойчивости находятся в одном сорте (Turska-10016), т.е. в ряду сцепления RT/RT, а гены восприимчивости - в другом сорте (Rio - rt/rt), то при скрещивании этих сортов в F2 появляется 81,94% устойчивых растений, будь то первый, второй или оба гена устойчивости (табл. 4.5). Из числа устойчивых 83,05% растений будут иметь два гена устойчивости (R и T), и среди них 21,74% будут гомозиготными по обоим генам устойчивости.
Если бы эти гены были независимыми, т.е. находились в различных хромосомах, то в F2 появилось бы 93,75% устойчивых растений, но из них только 56,25% содержали бы оба гена устойчивости и лишь 6,25% были бы гомозиготными по обоим генам (см. табл. 4.5).
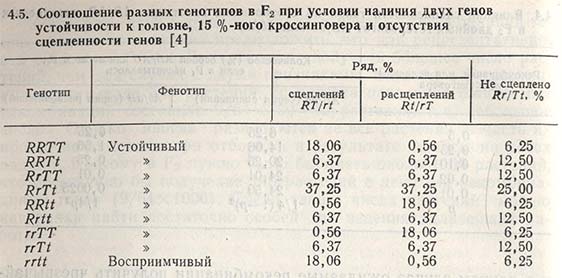
В случае когда первый ген устойчивости находится в одном сорте (kt/kt), а второй - в другом сорте (rT/rT), т.е. в ряду расщепления, число желаемых рекомбинаций изменяется. В F2 почти 99,5% растений будут устойчивы, 50,5% будут обладать обоими генами устойчивости, но только 0,56% растений будут гомозиготными по обоим генам устойчивости к головне пшеницы (см. табл. 4.5).
Данный пример показывает, в какой мере сцепленность генных локусов влияет на рекомбинацию полезных признаков; чтобы такую рекомбинацию получить и выделить отбором, необходимо в F2 иметь многочисленную популяцию растений с рядами расщепления, а не с рядами сцепления.