Методы отбора растений-самоопылителей в популяции, сформированной при гибридизации
Популяция, сформированная плановой гибридизацией различных сортов, представляет собой генетически наиболее ценную популяцию для проведения отбора и получения новых сортов. Ее отличие от естественных или местных популяций растений-самоопылителей заключается в следующем: отбор в гибридной популяции начинается не с гомозиготных растений, находящихся в популяции, а с выявления гомозиготных и гетерозиготных растений, которые в результате оплодотворения гамет растений F1 появляются в поколении F2.
В данном случае также применяют массовый и индивидуальный отборы или их комбинации. Однако в каждом новом поколении расщепление происходит иначе, чем в предшествующем и последующем, поэтому нужно анализировать каждое поколение, принимая при этом во внимание генетическую структуру особи, число выращиваемых растений и число отобранных генотипов, т.е. интенсивность отбора.
Метод отбора в смеси
Этот метод еще называют методом популяций или методом выращивания в смеси [bulk (англ.), Ramsch (нем.)]. Он был впервые введен в селекцию в начале текущего столетия Нильсоном-Эле (Швеция). Позднее метод отбора в смеси применяли и теоретически разработали на растениях-самоопылителях (ячмень, пшеница, овес, соя и т.д.) Харлан, Мартини и Стивенс в США и многие другие.
Рекомбинации генов в гетерозиготном состоянии у растений - самоопылителей использовать нельзя, так как процесс самооплодотворения приводит к гомозиготности, которая, начавшись в F2, постоянно увеличивается, достигая в F6 96,9 %, в F10 - 99,8 % (см. табл. 4.3), даже если материал выращивают целиком в смешанной массе до F6, и только тогда начинают отбор. Практически это выглядит следующим образом.
1) После скрещивания сортов А Х В, C X D и т.д. семена F1 от каждой комбинации скрещивания выращивают отдельно, чтобы получить достаточное количество семян для F2 (рис. 12.5).
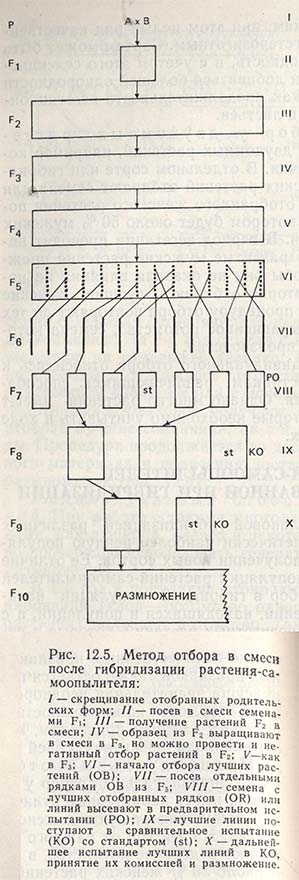
2) В F2 выращивают совместно несколько сотен или тысяч растений по одной комбинации скрещиваний, как указано в главе 11. Растения F2 от каждой комбинации скрещиваний обмолачивают вместе. Поскольку для получения F3 нельзя высевать все семена (на это потребовались бы огромные площади), берут только образец семян, принимая во внимание, что в каждой комбинации скрещиваний опять будет получено несколько сотен или тысяч растений в зависимости от вида. Эту же процедуру повторяют в F4 и F5, т.е. в поколениях, где расщепление признаков проявляется еще сильнее.
3) В F6, когда достигнутая по всем признакам гомозиготность составляет 96,9%, начинают индивидуальный отбор генотипов, удовлетворительных по своим признакам, т.е. тех, которые содержат желательные рекомбинации признаков родителей.
4) В F7 и последующих поколениях проводят отбор по потомству, выделяют лучшие линии, передают их в сравнительное испытание для оценки по урожайности и, наконец, приступают к созданию новых сортов (рис. 12.5).
На ранних этапах селекции некоторые авторы получали в смеси от F2 до F6 все комбинации скрещиваний, проведенные в течение одного года, и затем в этих смесях начинали индивидуальный отбор. Однако в настоящее время такой путь практикуется редко, ибо селекционеры стремятся уделять больше внимания отдельным комбинациям скрещиваний.
Преимущество метода отбора в смеси заключается в том, что получение огромного числа комбинаций скрещиваний отличается экономичностью, т.е. не требует больших затрат труда и средств на проведение наблюдений и отборов. Однако данный метод имеет и недостаток: при его использовании можно утратить или не заметить некоторые ценные генотипы, поскольку селекционер недостаточно эффективно использует селекционный материал. При этом особенно проигрывают молодые специалисты, которым необходимо лучше знать прорабатываемый материал и значение эффективности отбора в отдельных поколениях. О ценности метода отбора в смеси ведется дискуссия главным образом вокруг точки зрения на эффективность естественного отбора от F1 до F6, происходящего ввиду конкуренции между различными генотипами в популяции.
Поскольку полученная гибридная популяция-смесь состоит из генотипов, характеризующихся различной продуктивностью и различным взаимодействием с условиями окружающей среды, в ней закономерно происходит естественный отбор. Смысл этого отбора заключается в том, что одним генотипам он благоприятствует, в то время как другие генотипы исключаются и при этом сокращается частота их проявления. Если два генотипа находятся в состоянии конкуренции друг с другом, тогда их выживаемость зависит от:
- количества семян, производимых каждым генотипом;
- количества семян, дающих потомство.
Теоретическую кривую выживаемости ослабленного генотипа, по Алларду, можно рассчитать, используя следующую формулу:
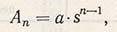
где Аn - количественное соотношение ослабленного генотипа в п поколениях; а - исходное соотношение ослабленного генотипа; s - селекционный индекс, учитывающий число полученных семян и численность потомства, полученного из этих семян.
Например, если два генотипа находились в смеси в одинаковом процентном соотношении 50:50 (A1 = 0,5), значение величины выживаемости для лучшего генотипа было равно 1, а для ослабленного генотипа - 0,9, тогда селекционный индекс равен 0,9. В таком случае ожидаемое соотношение, или репрезентативность ослабленного генотипа, например в F5, составит:
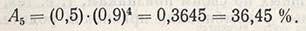
Это означает, что исходная 50%-ная репрезентативность ослабленного генотипа через пять поколений снизилась до 36,45%, а доля генотипа, давшего большее число семян и больше потомства, увеличилась до 63,55%.
Если генотипы, частота проявления которых возрастала на протяжении поколений, действительно являются родственными, то можно констатировать, что естественный отбор помог селекционеру. Сансон приводит результаты исследования эффективности естественного отбора, действовавшего в смеси на протяжении 28 поколений у 379 комбинаций скрещиваний сортов ячменя. Через 20 поколений вся популяция превзошла по урожайности стандартный сорт Atlas, чего не наблюдалось в более ранних поколениях. По мнению Сансона, называющего этот метод «эволюционным методом селекции растений», увеличение урожайности произошло потому, что в данной популяции естественный отбор благоприятствовал высокоурожайным генотипам. В конце концов, такие генотипы отбираются как чистые линии и их размножают как новый сорт.
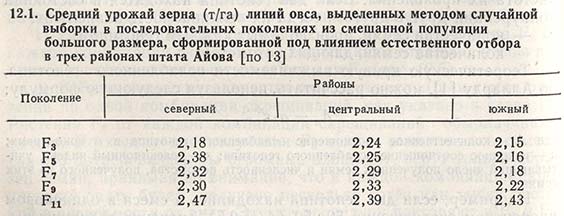
Фрей указывает, что Гонзалес в своей докторской диссертации приводит данные об изучении влияния естественного отбора на популяцию, состоявшую из смешанных семян F2 250 комбинаций скрещиваний сортов овса, от которых вплоть до F12 в трех географических районах штата Айова (США) брали образцы-смеси. Методом случайной выборки из поколений F3, F5, F7, F9 и F11 для каждого из трех географических районов были взяты 32 линии. Все линии анализировали в 15 питомниках, различавшихся плодородием почвы и дозами вносимых минеральных удобрений. Во всех районах было отмечено увеличение урожайности от F3 до F11 (табл. 12.1), что можно объяснить более высокой выживаемостью продуктивных генотипов.
В то же время некоторые примеры показывают, что эффективность естественного отбора в больших смешанных популяциях с селекционной точки зрения может оказаться вредной.
Дженнингс и Херрера рассчитали эффективность естественного отбора от F2 до F6 в популяции большого размера, полученной от скрещиваний двух сортов риса: высокостебельного Peta и низкостебельного TN1. Растения с площадью питания 30X30 см выращивали в вариантах без азота и на фоне высоких доз азота. В результате конкуренции между высокорослыми и низкорослыми генотипами популяции число растений с низкими стеблями из поколения в поколение уменьшалось, хотя теоретически ожидали, что в F6 оно составит 49% (табл. 12.2). Высокорослые растения при данной площади питания сильнее кустились и имели преимущество в конкуренции за свет. Однако в группе из 36 высокорослых и 36 низкорослых растений, отобранных в F6 методом случайной выборки и испытанных как чистые линии в питомнике с дозой 80 кг/га азота, линии с низкими стеблями в значительной мере превосходили по урожайности линии с высокими стеблями (табл. 12.3).

Наряду с этим эффективность естественного отбора в смешанной популяции как для различных условий, так и по желательным признакам может колебаться. Если сформировать большую популяцию и высеять растения на значительном расстоянии друг от друга (за счет чего снижается конкуренция между разными генотипами), то естественный отбор не приведет к большим изменениям в количественном составе генотипов. Селекционер, ставящий задачу усилить действие естественного отбора, должен просто создать условия для его проявления.

Улучшение метода отбора в смеси. Для повышения эффективности отбора в смеси прибегают к различным, вариантам его улучшения как по срокам, так и по способу вмешательства в популяцию.
Гибридные поколения, использование которых позволяет более быстро выделять новые линии, выращивают в смеси только до F4, но не до F6, что больше всего характерно для данного метода. В F4 начинают индивидуальный отбор соответствующих растений, а в дальнейших поколениях для изоляции новых линий применяют метод педигри. Такой путь требует меньше времени на получение новых линий: их создают за 1-2 поколения. В случае неотложной потребности в выведении новых сортов это может иметь большое значение для продвижения в производство данной культуры.
Позитивный или негативный отбор в смеси. В очень большой по численности популяции, не подвергшейся действию какого-бы то ни было вмешательства, средний образец, отбираемый из поколения в поколение, содержит значительное число генотипов, вообще не попадающих под влияние отбора, если его начинают в F5 или F6 (см. рис. 12.5). При селекции на низкорослость, скороспелость или позднеспелость и т.д. это могут быть, например, генотипы с высоким стеблем.
Чтобы отбираемый образец содержал больше растений с желаемыми комбинациями признаков и отбор был эффективным, можно, начиная с поколения F3, применить позитивный отбор. Его проводят по глазомерно легкоразличимым признакам. Например, отбирают все растения с низким и средненизким стеблем, обмолачивают в смеси и в следующем поколении выращивают также в смеси. При таком способе большинство растений с высокорослым стеблем выбраковывается.
При отборе на продолжительность вегетации легче применять негативный отбор. С наступлением сроков уборки урожая раннеспелые растения трудно отличить от позднеспелых. Поэтому в период, когда они легче всего различаются по скороспелости, поздно зацветающие растения удаляют прорывкой или срезают ножницами; все оставшиеся растения обмолачивают и в следующем поколении выращивают их в смеси.
Метод отбора в смеси с различными его вариантами применяют в настоящее время в гибридных поколениях для выведения сортов пшеницы, ячменя, овса, сои, вики, гороха и других видов-самоопылителей.
Метод педигри
Метод педигри представляет собой метод индивидуального отбора растений в расщепляющихся поколениях и прослеживания родословной отобранных растений вплоть до получения гомозиготных линий. Учитывая, что метод педигри применяют у растений-самоопылителей гораздо чаще, чем массовый отбор, он будет рассмотрен подробнее по отдельным поколениям (рис. 12.6).
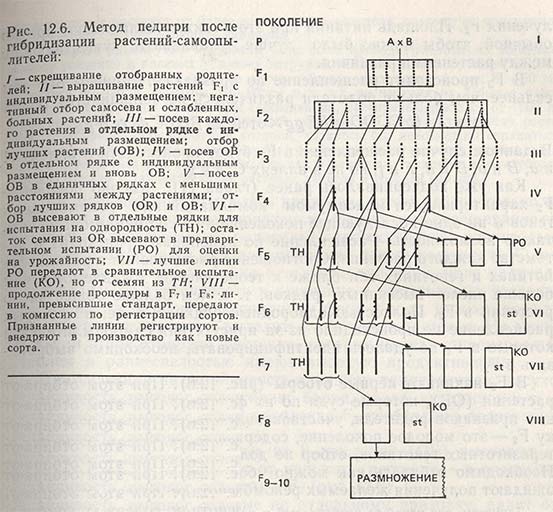
Поколение F1. Семена F1, полученные от скрещивания разных родителей, высевают в производственных условиях с большим, чем обычно, расстоянием между растениями и как можно большими междурядьями. Этим добиваются лучшего развития растений и получают большее количество семян. В F1 необходимо удалить негибридные растения, появившиеся от самооплодотворения после неполного или слишком поздно проходящего свободного переопыления. Растения-самоопылители (в действительности растения маточного сорта) можно идентифицировать, если отцовская форма является носителем некоторого доминантного гена (красная окраска, отсутствие опушенности и т.д.). Гибридные растения будут обнаруживать доминантные признаки отцовской формы, в то время как негибридные растения - признаки материнской формы (белая окраска, опушенность и т.д.). Если в гибридной комбинации материнский сорт был носителем доминантных признаков, то не в F1, а только в F2 появится возможность идентифицировать потенциальную особь-самоопылитель. В F1 можно выбраковать комбинации, оказавшиеся явно худшими.
Поколение F2. Семена от каждого растения F1 обмолачивают отдельно и высевают отдельными рядками или делянками для получения Площадь питания при этом также увеличивают против обычной, чтобы можно было лучше улавливать существующие между растениями различия. В F2 происходит расщепление по признакам, причем оно тем сильнее, чем больше родители различались по большинству генов:
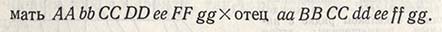
В данном случае расщепление в F2 будет происходить в аллеях A и а, В и b, D и d, F и f, но не в аллеях С, е и g.
Как уже подчеркивалось ранее, именно поколение F2 характеризуется максимумом комбинаций или рекомбинаций генов и ни одно последующее поколение не в состоянии обеспечить такую возможность. Расщепление по признакам происходит в потомстве каждого растения. Соотношение расщепления в разных фенотипах и генотипах тем ближе к теоретически ожидаемому, чем больше число высеянных рядков, т.е. чем больше общее число растений в F2. Полностью однородные рядки, т.е. те, в которых расщепление не происходило из-за присутствия самоопылителей и которые в F1 не удалось идентифицировать, необходимо выбраковать в F2.
В F2 начинают первые отборы (рис. 12.6). При этом отбирают растения (ОВ), которые судя по их фенотипу, содержат комбинации признаков родителя, участвовавшего в скрещивании. Поскольку F2 - это молодое поколение, содержащее пока только 50% гетерозиготных генотипов, отбор не должен быть излишне жестким. Необходимо отбирать как можно больше растений, от которых ожидают получения желаемых рекомбинаций генов.
Допустим, что какой-то низкорослый и позднеспелый сорт скрещивают с высокорослым и раннеспелым, надеясь получить при этом новый сорт, который был бы низкорослым и раннеспелым. Признак высоты стебля в F1 носит промежуточный характер наследования, что указывает на аддитивный эффект генов. В F2 происходит расщепление по признаку высоты стебля с колебанием от высокорослых растений (как у более высокорослого родителя) до низкорослых (как у более низкорослого родителя); именно поэтому наибольшее число растений оказывается промежуточным по данному признаку. Что касается продолжительности периода вегетации, то в F1 обнаруживается частичная доминантность по признаку раннеспелости, а в F2 - непрерывная изменчивость, как и по высоте стебля, однако расщепление идет активнее в направлении признака раннеспелости.
В данном случае прежде всего нужно отбирать низкорослые и раннеспелые растения, так как этого требует рекомбинация родительских признаков. Таких растений в F2 мало, поэтому полезно отбирать также средневысокорослые и раннеспелые, поскольку у части растений со средневысокорослыми стеблями в F2 произойдет расщепление и появятся низкорослые особи. Можно проводить отбор и средневысокорослых и среднеранних растений, ожидая, что в последующих поколениях при расщеплении появится некоторое количество низкорослых и раннеспелых особей. Не следует отбирать высокорослые и позднеспелые растения, так как при аддитивном действии генов, ответственных за признак высоты стебля, и большинстве доминантных генов, ответственных за признак раннеспелости, в последующих поколениях таких растений едва ли появятся со значительной частотой низкорослые и раннеспелые растения.
Принимая во внимание, что оба признака имеют полигенный характер наследования, в F2 можно ожидать также трансгрессивного расщепления. Особый интерес представляли бы низкорослые растения от более низкорослого родителя и раннеспелые растения от более раннеспелого родителя, поэтому на присутствие таких особей следует обращать внимание.
Отбор по упомянутым признакам чаще всего проводят глазомерно. Измерить высоту растений в полевых условиях нетрудно, если же высота растений задана, то низкорослые растения намного легче отбирать глазомерно (у сои 40-50 см, v ячменя 60-70, у пшеницы 70-80 см и т.д.), не измеряя их. Обычно отбор проводят не по одной комбинации скрещиваний, насчитывающей в F2 от 1000 до 3000 растений, а по сотням комбинаций скрещиваний с десятками тысяч растений, т.е. проделывая работу, с которой, казалось бы, невозможно справиться. При выявлении признака раннеспелости соответствующие растения удобно отмечать в период цветения этикетками, так как у некоторых видов во время уборки урожая трудно установить, какие растения зацвели и созрели раньше.
Для большей простоты здесь приведена схема отбора только по двум рекомбинированным признакам. Между тем скрещивание генетически дивергентных родителей предоставляет значительно большие возможности для рекомбинаций генов; поэтому в F2 следует обращать внимание и на остальные признаки. Если родители различались по устойчивости к ржавчине в естественных условиях или на искусственном инфекционном фоне в поле, нужно провести оценку на этот признак и выделить устойчивые растения. На завершающем этапе отбирают растения, которые наряду с низкорослым стеблем и раннеспелостью имеют высокую продуктивность.
При селекции на устойчивость к низким температурам, засухоустойчивость, высокое содержание белка, масла, витаминов и т.д. растения из F2 до посева на поколение F3 необходимо подвергнуть лабораторному анализу.
Поколение F3. Отобранные из F2 растения (ОВ) высевают по методу педигри, т.е. семена от каждого растения - в отдельном рядке с площадью питания, как в F2 (рис. 12.6). Растения F2, гомозиготные по отдельному признаку, дадут однородное по этому признаку потомство, в то время как по другим признакам может произойти расщепление. Например, в F3 потомство всех растений будет иметь низкорослые стебли, но и различаться по раннеспелости, как и потомство, выравненное по признаку раннеспелости, но расщепляющееся по высоте стебля.
F3 - первое поколение после гибридизации растений-самоопылителей, в котором начинают формирование линий. Поэтому отбор в F3 основан не только на оценке признаков отдельных растений, но и на оценке отдельных потомств. В F3 проводят отбор растений, обладающих желательными рекомбинациями признаков, причем прежде всего из потомства, в наибольшей мере отвечающего поставленной цели, а также из потомства, в целом не удовлетворяющего требованиям, так как в нем всегда есть отдельные растения с Ценными признаками.
Поколение F4. Отобранные в F3 растения (ОВ), высевают в F4 по методу педигри, т.е. каждое растение в отдельный рядок. В этом поколении гомозиготность составляет 87,5%, т.е. большинство потомства обнаруживает выравненность по отбираемым признакам, и здесь уже можно говорить о формировании линий. Поэтому отбор в F4 большей частью основан на отборе потомства и в меньшей мере - на отборе отдельных растений (рис. 12.6).
Потомства, т.е. линии, которые по своим признакам в наибольшей мере отвечают поставленным целям и, согласно глазомерной оценке, отличаются выравненностью, отбирают для посева в F5. Так как отобранное потомство еще не совсем гомозиготно по некоторым признакам, целесообразно перед уборкой урожая отобранных рядков (OR) провести индивидуальный отбор приблизительно 5-10% растений, которые в F5 будут высеяны по методу педигри как тест на однородность или гомозиготность (ТН). Остальные растения с отобранных рядков убирают вместе, семена используют для проведения испытания на урожайность в F5. В оставшихся потомствах, где еще происходит расщепление по признакам, индивидуально отбирают лучшие растения (ОВ).
Начиная с F2, наряду с отбором по отдельным признакам постоянно идет отбор на урожайность: отбирают лучшие и наиболее продуктивные растения; однако без прямого испытания на урожайность объективные данные о продуктивности отдельных растений и линий получить нельзя. Чтобы параллельно с глазомерной оценкой получить как можно больше объективных данных об основных агрономических признаках отобранных потомств или линий, с каждого отобранного рядка (OR) берут небольшое количество семян для испытания на зимостойкость, качество, устойчивость к болезням и т.д.
Поколение F5. Семена из отобранных потомств или линий F4 высевают для предварительного или подготовительного испытания (РО) на урожайность (рис. 12.6). Линии испытывают на урожайность, сравнивая их со стандартным сортом в двух или более повторностях в зависимости от количества имеющихся семян. При этом необходимо, чтобы способ и густота посева, дозы вносимых минеральных удобрений и другие агротехнические приемы были близки к применяемым в производстве.
На засеянных делянках в тесте на гомозиготность (ТН) от каждой находящейся в предварительном испытании (РО) линии отдельно выращивают растения с индивидуальным размещением. На тех же делянках находятся и рядки отобранных в F4 растений, которые еще не имеют своих линий в предварительном испытании.
В полевых условиях и на первом, и на втором материале проводят фенологические наблюдения, оценки на раннеспелость, устойчивость к болезням, к полеганию и т.д. В F5 прорабатываемый материал уже значительно выравнен, так как в этом поколении гомозиготность достигает 93,8%. Как правило, все рядки растений, отобранных предварительно из одного ряда, не совсем одинаковы и в F4 самым необычным образом реагируют на тест гомозиготности (ТН), чего нельзя заметить на делянках линий, находящихся в предварительном испытании (РО).
В этом же году завершаются все анализы образцов, которые берут в линиях, находящихся в предварительном испытании. На основе этих сведений, полевых наблюдений и данных урожайности выносится решение о том, какие линии из предварительного испытания поступят в сравнительное испытание в F6 и какие будут исключены из дальнейшей работы. Поскольку на этом этапе от каждой линии уже получено несколько килограммов семян, можно провести подробные анализы качества урожая, испытание на устойчивость к болезням и другие необходимые исследования.
Из потомств отобранных в F4 растений (ОВ), которые в F5 не были включены в предварительное испытание, отбирают лучшие для формирования новых линий в новом предварительном испытании, а также растения для теста на гомозиготность (TH).
Поколение F6. Лучшие линии из предварительного испытания передают в сравнительное испытание (КО) для оценки на урожайность. Следует подчеркнуть, что семена для сравнительного испытания берут не из предварительного испытания (из-за возможности механического засорения во время уборки и обмолота и из-за недостаточной выравненности), а из теста на гомозиготность (ТН) данных линий (рис. 12.6). Если в ТН находилось 10-20 рядков длиной 1-2 м, то можно получить достаточное количество семян для посева этих линий в питомнике сравнительного испытания в пяти повторностях. В сравнительное испытание нужно включать один или два сорта-стандарта и применять агротехнику возделывания, сходную с принятой в производственных условиях.
Наряду со сравнительным испытанием в F6 закладывают и питомник предварительного испытания вновь отобранных линий, проводя посев по той же методике, что и в поколении F5. Потомства отобранных в F5 растений выращивают на отдельных делянках. Здесь располагаются защитные рядки линий ТН находящихся в сравнительном испытании, рядки линий ТН, находящихся в предварительном испытании, и рядки отобранных растений из потомств, которые из-за расщепления не попали в предварительное испытание.
На основе наблюдений, проведенных в полевых условиях, лабораториях и теплицах, а также анализов полученного в испытаниях урожая для дальнейшей работы отбирают лучшие линии. Данные урожайности достоверны только в том случае, если они получены по результатам сравнительного испытания, питомники которого закладывают в большом числе повторностей рендомизированными блоками. Линии с лучшей урожайностью и другими ценными признаками поступают в сравнительное испытание в F7. В оставшемся материале отбирают новые линии и растения для предварительного испытания, тестов на гомозиготность и формирования новых линий.
Поколения F7-F10. С целью подборки большего числа хороших линий в F7 и последующих поколениях весь процесс отбора повторяют точно таким же образом, как и в F6. Лучшие линии испытывают в сравнительном испытании (F6-F8) в среднем около трех лет. На основе результатов этого испытания несколько лучших линий передают для регистрации в качестве новых сортов. В Югославии комиссия по регистрации сортов проводит испытание новых сортов однолетних растений-самоопылителей три года в зависимости от распространенности культуры (пшеница - в 20 районах, ячмень - в 11, соя - в 9 и т.д.) в большинстве географических районов страны. Питомники испытаний закладывают в пяти повторностях рендомизированными блоками с одним или двумя стандартами, которыми служат сорта, являющиеся в настоящее время ведущими в стране по продуктивности и качеству урожая. Новые сорта оценивают на основе результатов трехлетнего изучения. Сорта, превосходящие по урожайности стандарт Или одинаковые с ним, но имеющие лучшее качество или какие-то другие важные признаки, регистрируются как новые. Такие сорта заносят в каталог зарегистрированных сортов, они могут быть использованы в производстве. Учреждения-оригинаторы вновь зарегистрированных сортов совместно с семеноводческими организациями заботятся о дальнейшем их размножении и внедрении.
Такова вкратце общая характеристика метода отбора педигри, который фактически является методом непрерывного индивидуального отбора после гибридизации. Отбор в основе своей проводят не только по фенотипу, но и по генотипу, поэтому применение данного метода оказывается весьма успешным при выделении лучших линий в расщепляющемся поколении.
При создании новых сортов пшеницы, ячменя, сои, гороха, томатов, перца овощного и других видов растений-самоопылителей в основном применяют метод педигри в гибридных поколениях. Югославские высокоурожайные сорта пшеницы (Sava, Zlatha dolina, Biserka, Novosadska rana 1 и Novosadska rana 2, Partizanka, Kragujevacka-56 и многие другие) и ячменя (Mursa, Novosadski-27, Partizan и др.) выведены методом педигри.
Однако этот метод имеет недостатки, и основной из них - это большие затраты труда и рабочей силы. При работе этим методом необходимо проводить индивидуальный отбор растений или колосьев, просмотр и индивидуальный обмолот растений, индивидуальный посев, индивидуальную отметку отобранных растений и линий и т.д. На это уходит немало усилий, особенно когда за год необходимо провести более ста комбинаций скрещиваний. Второй главный недостаток метода педигри заключается в постоянном отборе одинаковой доли гетерозигот, которые и без вмешательства человека достигли бы гомозиготного состояния в поздних поколениях, как это происходит при использовании метода отбора в смеси. Наряду с этим эффект гетерозиса и доминантность довольно долго маскируют генотипическую ценность отобранных растений, что снижает эффективность отбора. Данный недостаток компенсируется отбором гомозигот в ранних поколениях и ускоренным получением линий для испытания на урожайность, а это возможно при использовании метода отбора в смеси. Отличие метода педигри от метода отбора в смеси состоит в том, что при использовании в первом случае конкуренция между различными генотипами сведена до минимума и в основном не влияет на результаты отбора.
Ввиду перечисленных недостатков, применение находят и некоторые варианты метода педигри.
Метод педигри с прерывающимся индивидуальным отбором
Этот метод, рекомендованный Акерманом и Мак Кеем в Швеции и Лаптоном и Уайтхаузом в Англии, называют методом потомства F2. Заключается он в следующем. В F2 проводят индивидуальный отбор отобранных растений (OR). В F3 потомство каждого отобранного растения выращивают в отдельном ряду. В F4 каждое потомство без применения отбора испытывают по урожайности в питомнике предварительного испытания (РО), а в F5 и F6 испытывают потомства (без применения отбора) в питомнике сравнительного испытания (КО), чтобы собрать сведения об урожайности. Потомства с наивысшей урожайностью и некоторыми другими ценными признаками отбирают для дальнейшей работы (рис. 12.7).
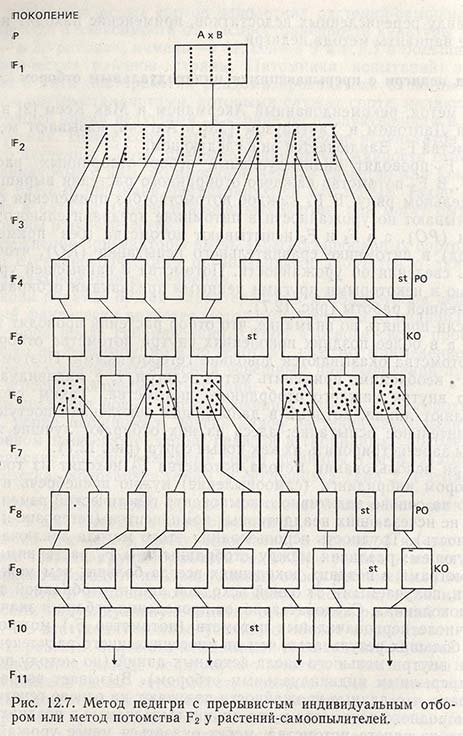
Если принять во внимание, что отбор растений проводят только в F2, а в более поздних поколениях внутри потомства отбора нет, то потомства оказываются довольно гетерогенными. Поэтому в F7 или F8 необходимо применить метод педигри, т.е. индивидуальный отбор внутри каждого отобранного потомства. Таким способом выделяют линии, которые в дальнейших поколениях поступают в сравнительное испытание; здесь из них отбирают лучшие линии, чтобы зарегистрировать их как новые сорта (рис. 12.7).
При использовании метода потомства F2 исходят из того, что фактором инбридинга (самоопыление) нужно пренебречь и вести отбор на основе аддитивного компонента генетической изменчивости, а не исчезающих неаддитивных компонентов (гетерозис и доминантность). Трудность использования этого метода заключается в следующем: различия между отобранными в F2 растениями и их потомствами в поздних поколениях всегда больше, чем между линиями, получаемыми от одной исходной линии, отобранной в поздних поколениях. Следовательно, от проведения отбора в значительном числе первоначальных потомств (потомство F2) можно ожидать больших результатов, чем от многочисленного расчленения на линии внутри меньшего числа исходных линий (по методу педигри с непрерывным индивидуальным отбором). Вызывает возражение тот факт, что данные урожайности получают на основе генетически неоднородного потомства и что линии, выделяемые в поздних поколениях из одного потомства, могут оказаться менее урожайными, однако в высокоурожайных потомствах присутствует большее число лучших генотипов, чем в низкоурожайных.
Все эти рассуждения обоснованы генетически, и метод потомства F2 можно было бы отнести к самым результативным методам отбора, проводимым после гибридизации растений-самоопылителей, если бы он не был обременен практическими трудностями.
Вообще говоря, чтобы не слишком сужать во время отбора Масштабы генетической изменчивости, необходимо из F2 отбирать не менее 100 растений. При известной выбраковке целых потомств в сравнительном испытании из этого количества останется около 70 растений на комбинацию. Если в селекционную программу ежегодно включать 100 комбинаций скрещиваний, то только в сравнительном испытании от одного цикла скрещиваний участвовало бы 7000 потомств плюс столько же из предварительного испытания и предыдущих лет, а это работа, с объемом которой справиться физически невозможно.
Поэтому метод потомства F2, или метод педигри с прерывающимся индивидуальным отбором, можно успешно применять только на одном этапе комбинации скрещиваний или в учреждениях, где проводят ограниченное число скрещиваний.
Метод односемянного потомства
С целью ускорения отбора и упрощения его методов используют разные модификации метода педигри, из которых особенно следует упомянуть метод односемянного потомства (single-seed descent method). Он был предложен Гоулденом и позже применен в селекции сои.
Метод односемянного потомства ставит задачей испытание потомства каждого растения F2. Поскольку изучаемый материал, полученный от всех семян каждого растения F2, использовать невозможно, в каждом следующем поколении от каждого растения методом случайной выборки берут только одно семя (рис. 12.8). При работе с соей из практических соображений берут один боб, а иногда 2-3 семени. Лучшие растения отбирают, добившись достаточной степени выравненное™ (гомозиготности), что обычно наблюдается в F5. В F6 семена от каждого отобранного растения высевают в отдельном рядке. Если это уже выравненные линии, единицей отбора становится целый рядок. В F7 отобранные линии, испытывают на урожайность, проводят отбор наиболее урожайных линий и тех, которые соответствуют остальным признакам (содержание белка и масла, устойчивость к болезням и т.д.). В дальнейших поколениях индивидуальный отбор продолжают (рис. 12.8).
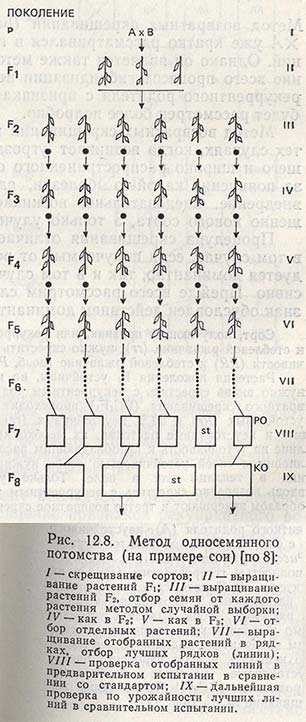
Используя этот метод, отбор можно ускорить во времени. Поскольку для получения следующего поколения от одного растения берут только одно семя, изучаемый материал можно выращивать в теплице и при соответствующем регулировании освещенности и температуры получать два-три поколения за один год.
Преимущества метода односемянного потомства многочисленны, но есть и существенные недостатки. Главная опасность заключается в том, что при отборе одного семени с растения не только охватывается лишь часть всех лучших генотипов, но и выделяется та часть растений F2, которая не содержит желательных рекомбинаций генов. В зависимости от числа растений в F4, как и в F2, наступает генетический дрейф, вследствие которого неизбежны значительные потери ценных генов, особенно если родители различаются по большому числу генов. Далее, посев растений на большем расстоянии друг от друга и в теплицах не позволяет проводить оценку на устойчивость к полеганию и другие признаки; из-за этого позже снижается эффективность отбора в полевых условиях.