Три концепции подбора родительских пар
Существуют три концепции подбора родительских пар для скрещивания: концепция сорта, концепция признака и концепция гена.
Концепция сорта означает подбор большого числа сортов для скрещивания, если предположить, что в новых сортах будет получена желаемая комбинация положительных признаков. Эта концепция характерна для ранних этапов селекции растений путем гибридизации, когда было мало известно о генетике признаков, подлежавших передаче от разных родителей. В настоящее время концепция сорта не находит широкого применения в прогрессивных селекционных программах, но все же принимается в учет, особенно когда приходится переходить на новую программу или когда из-за недостатка времени не удается подробно ознакомиться с признаками сортов, находящихся в распоряжении селекционера. Успех этой концепции определяется в основном использованием большого числа комбинаций скрещиваний, хотя некоторые из них могут дать перспективные линии и новые сорта. В подобной программе много риска и неопределенности, об этом говорит выбраковка огромного числа целых комбинаций скрещиваний, на проведение которых уже затрачено немало труда и времени. Поэтому более успешным оказывается использование концепции признака.
Концепция признака. Под этой концепцией подразумевается подбор родительских пар для скрещивания на основе знания признаков родителей, которые хотят объединить в новых сортах. Существуют фенотипически различные сорта, которые могут иметь сходную или одинаковую зимостойкость, скороспелость, высоту стебля, массу зерен и т.д. Отбор родителей на основе данной концепции требует поиска различий между сортами в признаках, которые должны быть объединены. Поэтому скрещивания базируются на очень большой генетической дивергентности, существующей между родителями, и целенаправленном планировании комбинаций скрещиваний. Таким образом возникает возможность сокращать число комбинаций скрещиваний (в сравнении с концепцией сорта) и возрастают шансы на успех в получении перспективных линий и новых сортов. В комбинации скрещивания, т.е. при одинаковом генетическом участии обоих родителей, следует учитывать и тот факт, что каждый из них содержит или больше положительных, или меньше отрицательных признаков.
Если селекция ведется методом полного возвратного скрещивания, следует в основном добиваться большего числа положительных признаков не у отдающего их родителя (донора), а у принимающего (реципиента, или рекуррентного родителя). По этой причине данный принцип подбора родительских пар в настоящее время применяют почти во всех программах создания новых сортов.
Концепция гена означает подбор родительских пар для скрещиваний на основе знания генетической структуры признаков, по которым ведется селекция. Проведенные до последнего времени исследования позволили определить генетическую основу ряда признаков у разных сортов и видов растений. Например, установлено, что устойчивость к стеблевой ржавчине пшеницы (Puccinia graminis tritici) обусловлена более чем 15 главными генами (Sr1, Sr2 и др.), к листовой ржавчине (P. recondita) - приблизительно 25 главными генами (Lr1 Lr2 и др.). Одни сорта содержат только один ген, другие - комбинацию Lr1 и Lr3, третьи - только Lr9 (дающий самый сильный эффект) и т.д.
То же обнаружено в отношении устойчивости к гельминтоспориозу (Helmintosporium ssp.) у ячменя, кукурузы, сорго и к другим патогенным организмам. Выявлена также генетическая основа устойчивости к ряду видов патогенных организмов у сахарной свеклы, томатов, бобов и других культур.
Озимость, например, представляет собой в значительной степени комплексный признак, включающий и устойчивость к низким температурам, и обязательно продолжительную яровизацию и т.д. Установлено, что признак озимости контролируется главными и малыми генами, которые могут обладать доминатным или аддитивным эффектом. Так, методом моносомных серий удалось обнаружить хромосомы пшеницы, являющиеся носителями генов озимости. Особое значение имеет тот факт, что некоторые озимые сорта могут содержать гены яровости и, наоборот, яровые сорта могут нести гены озимости.
Если принять во внимание полиплоидную природу пшеницы, овса и других культур, то неудивительно, что при скрещиваниях озимых сортов получают яровые, а при скрещиваниях яровых - озимые формы. Предположим, что озимость обусловлена только одним главным геном в одной единственной хромосоме каждого генома пшеницы (Triticum aestivus ssp. vulgare), таким образом, при скрещивании двух сортов
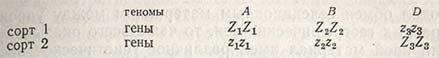
В случае полного доминирования генов озимости (ZZ) оба сорта будут иметь одинаковую степень озимости, растения F1 будут озимыми, а в F2 появится известный процент яровых растений (z1z1 z2z2 z3z3). При аддитивном эффекте генов сорт 1 будет иметь большую устойчивость к перезимовке, чем сорт 2, а в F2 произойдет трансгрессивное расщепление в обоих направлениях и появятся устойчивые генотипы от обоих родителей (Z1Z1 Z2Z2 Z3Z3), а также яровые генотипы (z1z1 z2z2 z3z3).
Если допустить, что: 1 - какой-то единичный признак обусловлен большим числом малых генов (характерно для количественных признаков); 2 - ген Z в геноме А дает иной эффект, чем в геноме В или D; 3 - один ген может быть модификатором или супрессором другого гена, то возникают большие возможности для проявления различий только по одному признаку между разными сортами и для комбинирования генов в потомстве.
Следовательно, чем лучше познаны эти различия, тем глубже удается селекционеру проникнуть в генетическую сущность некоторых признаков, а это позволит провести отвечающий требованиям подбор сортов-носителей данных признаков и осуществить более направленный подбор родительских пар для скрещивания, что, несомненно, приведет к успеху. Известный австралийский генетик и селекционер Френкель подчеркивал уже в послевоенные годы, что селекция больше не может опираться на наблюдения, а должна и обязана управлять генами количественных признаков.
Поэтому там, где это возможно, подбор родительских пар для скрещивания нужно проводить на основе концепции гена. Это достаточно хорошо известно в отношении признаков, обусловленных действием главных генов. Между тем существует немало количественных признаков с еще недостаточно изученным генетическим строением, тем более признаков различных сортов (число зерен, размеры плодов, содержание белка), обусловленных действием большого числа малых генов. В таком случае концепция признака представляется наиболее надежным и доступным средством подбора родительских пар.
В связи с подбором родителей для скрещивания нужно сказать несколько слов о принципе географической удаленности родительских пар. Этот принцип, в частности, отстаивал и применял советский селекционер-плодовод И. В. Мичурин, который считал, что лучшие новые сорта дает скрещивание или прививка географически удаленных сортов или даже видов. Позже эту идею поддерживали исключительно селекционеры по полевым культурам.
Понятие географической удаленности с начала текущего столетия и вплоть до второй мировой войны фактически представляло понятие генетической дивергентности. В различных государствах и институтах создавались многие сорта растений, и когда при этом происходил обмен селекционным материалом между учреждениями из разных географических зон, то чаще всего оказывалось, что тот или иной материал имел различное генетическое происхождение.
После второй мировой войны, когда развитие торговли, средств сообщения, открытие границ между государствами и т.д. привели к такому обмену информацией и материалом, как никогда ранее, географическая удаленность сортов и другого растительного материала утратила свой первоначальный смысл.
Сорт пшеницы Sava не обладает признаками, характерными для сортов Fortunato и Redcoat (см. табл. 7.3), не из-за их географической удаленности, а из-за наличия генетических различий между последними. Итальянский сорт Fortunato не имеет каких-либо типичных «географических» признаков, так как был получен при скрещивании японского сорта Akagomughi и голландского сорта Wilhelmina Tarve. Аналогично обстоит дело и с родословной сорта Redcoat (см. табл. 7.3). Некоторый географический отпечаток накладывает место, где проводится отбор, но при этом не происходит передача генов зимостойкости, если их не было ни у одного из скрещиваемых родителей. В то же время в любом месте можно создавать самые различные генотипы в зависимости от генетической дивергентности скрещиваемых родителей и направления отбора, осуществляемого селекционером.
Поэтому в настоящее время понятием генетическая дивергентность при подборе родительских пар пользоваться удобнее, чем понятием географическая удаленность. Географическая удаленность имеет свое значение при проявлении действия естественного отбора.
Стоит сделать упор и на то, что подбор сортов важен не только в программе гибридизации, но и при осуществлении какого-либо другого этапа, например когда в селекционный материал нужно внести некоторые изменения. При выполнении программы индуцирования мутаций с помощью облучения, химических препаратов или других средств также важно отобрать соответствующий материал. Можно ожидать, что он даст индуцированные мутации, а в сочетании с остальными признаками представит интерес и для генетических исследований или селекции растений.