Генетические основы селекции растений-перекрестников
Огромное число видов растений размножается с помощью системы перекрестного опыления. Перекрестное опыление, или аллогамия, - это такая система размножения, при которой мужские половые клетки одного растения оплодотворяют женские половые клетки другого растения. Оплодотворение происходит при помощи ветра или насекомых, а в отдельных случаях - в водной среде. Одни виды растений, как и животные, характеризуются полным разделением полов, у других генеративные органы разделены на одном и том же растении, третьи имеют двуполые цветки и генетические механизмы, предотвращающие самоопыление. Указанные различия обусловливают существование видов с развитым перекрестным опылением, а также видов, которые могут сохранять и систему самоопыления. К группе наиважнейших перекрестноопыляемых сельскохозяйственных культур относятся:

Из приведенного перечня видно, что в ходе эволюции выработавшееся у ряда видов перекрестное опыление привело к возникновению на одном растении мужских и женских цветков (обоеполые растения) или даже к появлению женских и мужских растений (раздельнополые растения). И все же значительное число видов имеет обоеполые цветки, причем перекрестное опыление полностью гарантировано или поддерживается наравне с самоопылением. У таких видов выработались особые механизмы генетической, морфологической и физиологической природы, которые привели к возникновению несовместимости, т.е. к неспособности развития семян после слияния мужской и женской гамет в одном и том же цветке, что способствует проявлению автостерильности.
Следует подчеркнуть, что с расцветом генетических исследований и разработкой новых методов селекции у ряда видов, раньше считавшихся строгими перекрестниками (подсолнечник, рожь и др.), выявлена возможность проведения принудительного самоопыления, а затем на его основе успешного создания инбредных линий.
Несовместимость
Для обеспечения перекрестного опыления, особенно у обоеполых цветков, в ходе эволюции возникли различные барьеры, препятствующие самоопылению. Существует несколько причин автостерильности или несовместимости, к которым относятся:
протандрия - когда в одном и том же цветке тычинки с пыльниками созревают раньше пестиков и пыльца попадает не на рыльца данного пестика, а на различные другие цветки;
протогиния - когда пестик созревает раньше тычинок с пыльниками, что также не может привести к оплодотворению в одном и том же цветке;
геркогамия - разделение на мужские и женские генеративные органы, при котором на одном и том же растении не может произойти самооплодотворение.
Наряду с этими морфологическими и механическими механизмами, обусловливающими перекрестное оплодотворение, причины несовместимости у черешни, огромного числа сортов яблони, груши и миндаля, сливы, вишни и маслины и далее у ржи, сахарной свеклы, люцерны, некоторых видов кормовых злаковых трав, табака и т.д. имеют генетическую и физиологическую природу. Льюис разделяет несовместимость у растений на две группы - гетероморфную и гомоморфную.
Гетероморфная несовместимость
Система несовместимости, основанная на различии в длине тычинок и пестика, называется гетеростилией. Тип цветка с длинным пестиком и пыльниками на коротких тычиночных нитях Льюис назвал pin (гвоздь, игла); тип цветка с коротким пестиком и длинными тычиночными нитями был назван thrum (сережка, букет).
Генетическая конституция типа pin - ss, типа thrum - Ss, при этом 5 полностью доминирует над s (рис. 5.1).
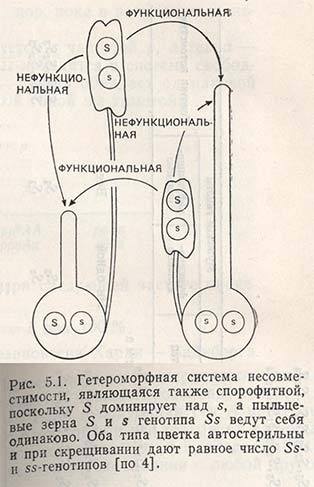
Оплодотворение происходит только между следующими генотипами:
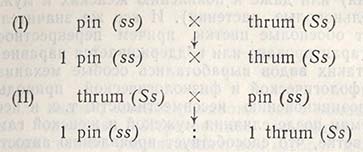
Комбинации pin X pin и thrum X thrum несовместимы, т.е. генотипы типа SS не возникают (рис. 5.1).
Гомоморфная несовместимость
Гомоморфная несовместимость разделяется на гаметофитную и спорофитную.
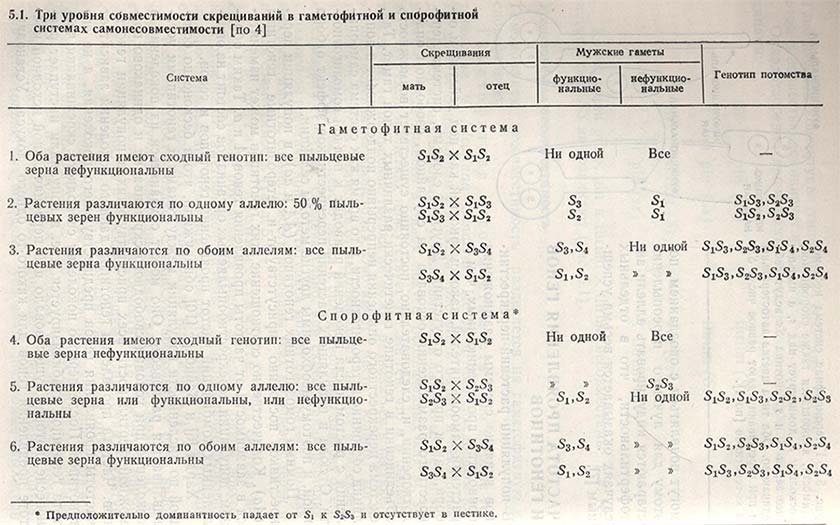
Гаметофитная несовместимость была впервые установлена у Nicotiana sanderae Истом и Мангельсдорфом. Позднее ее обнаружили у клевера красного и белого и некоторых других кормовых бобовых. Функциональность пыльцевых зерен зависит от ряда аллелей в одном локусе, обозначенных как s1, s2, s3, ... sn. Если в клетках присутствует тот же ген, что и в пыльцевом зерне, то пыльцевая трубка не может развиваться в пестике и оплодотворения не происходит. Оно имеет место только, когда пыльца несет гены, отличные от находящихся в пестике и яйцеклетке. Способность пыльцевого зерна к оплодотворению зависит от его собственного генотипа. Следовательно, при скрещивании растений с различной генетической конституцией может развиваться: полная несовместимость, совместимость половины всех пыльцевых зерен и полная совместимость (табл. 5.1).
Спорофитная несовместимость. Эта система несовместимости также обусловлена рядом аллелей в одном локусе, по в отличие от гаметофитной системы в данном случае функциональность пыльцевых зерен определяется генетической конституцией растения, которое их продуцировало (см. табл. 5.1).
Несовместимость создает сложности, поскольку у ряда видов невозможно получить инбредные линии для использования их на гетерозисной основе. И все же несовместимость стремятся применять главным образом в селекции овощных и цветочных культур. Об этом пойдет речь в главе, посвященной методам отбора.
Особенно надлежит учитывать явление несовместимости при возделывании овощных культур. Оно может обусловливать слабое оплодотворение и приводить к снижению урожаев, следовательно, нужно обязательно добиваться хорошего переопыления сортов. При неблагоприятных погодных условиях насекомые-опылители могут появляться с опозданием, поэтому для лучшего переопыления стараются индуцировать аллели автофертильности, что в отдельных случаях оказывается весьма успешным.
Частота проявления генов и генотипов
В популяции растений-перекрестников влияние системы размножения приводит к проявлению иных, чем у растений-самоопылителей, закономерностей как под действием отбора, так и при его отсутствии. Другими словами, система перекрестного размножения основана на случайном, свободном оплодотворении, и, следовательно, постоянно существует возможность объединения в генотипе генетически различных типов гамет. Так, система перекрестного опыления непрерывно поддерживает в популяции состояние гетерозиготности, в то время как система самоопыления с ростом числа поколений приводит к полной гомозиготности. Отсюда в популяциях растений-перекрестников эффект отбора иной, и поэтому необходимы другие методы селекции.
Помимо гомозиготных генотипов (AA и аа), в популяции перекрестников постоянно присутствуют и гетерозиготные генотипы (Aа). Количественно соотношение этих генотипов может изменяться в зависимости от частоты проявления генов А и а. Таким образом, популяция перекрестноопыляемых организмов состоит из определенных комбинаций частот проявления генотипов и генов.
Харди и Вайнберг обнаружили, что в бесконечно большой по численности популяции между генами и генотипами устанавливается равновесие. Оно может сохраняться из поколения в поколение при условии свободного скрещивания и одинаково возможной фертильности всех присутствующих в популяции генотипов. Частота проявления генотипов данного поколения зависит от частоты проявления генов предшествующего поколения, а не его генотипов. После первого поколения свободного скрещивания частота генотипов стабилизируется, т.е. в популяции наступает равновесие, которое существовало при первоначальной исходной частоте проявления генов для какого-то одного локуса. Установившееся равновесие сохраняется до тех пор, пока в популяции не изменится частота проявления генов.
Если в популяции ген А встречается с частотой р, а ген a - с частотой q и эти гены и их частоты находятся в системе свободного скрещивания, т.е. когда любая гамета обладает одинаковой вероятностью слияния с любой другой такой же гаметой:
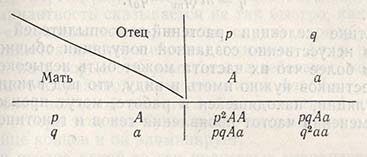
в популяции наступает равновесие при следующей частоте проявления генов и генотипов:
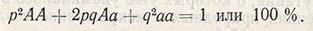
Это обобщенная формула закона равновесия Харди-Вайнберга.
Отличие частоты проявления генотипов от таковой в первоначальной популяции означает, что исходная популяция не находилась в состоянии равновесия. В системе свободного скрещивания с повышением частоты проявления генов равновесие частоты генотипов устанавливается в течение одного поколения независимо от частоты проявления генотипов у родителя.
Изменение частоты проявления генов в популяции
Бесконечно большая популяция - относительно устойчивая система, и все же состояние ее равновесия больше исключение, чем правило. Объясняется это тем, что на такую популяцию постоянно влияют факторы, нарушающие равновесие. К самым важным факторам, вызывающим изменение частоты проявления генов и генотипов, относятся миграции, мутации и отбор, которые могут действовать независимо друг от друга или чаще всего совместно.
Миграции
Миграция - включение новых особей в уже существующую старую популяцию. У растений, неподвижно связанных с местом обитания, это выражено в меньшей степени, чем у животных; в наибольшей же мере миграции характерны для насекомых, отличающихся подвижностью. Если семена растений разносятся на большие расстояния ветром и животными, то в старой популяции появляются новые мигранты. Миграцией можно считать также перенос пыльцы при помощи ветра и насекомых из одной популяции в другую, что в действительности есть миграция гамет (точнее генов) и появление в старой популяции новых генотипов.
Изменение частоты проявления гена (q) в новой смешанной популяции зависит от числа мигрантов (m) и различий между частотой проявления гена-мигранта (qm) и гена-хозяина (qo):
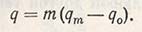
В практике селекции растений-самоопылителей возможные мигранты в искусственно созданной популяции обычно не учитываются, тем более что их частота может быть невысокой. У растений-перекрестников нужно иметь в виду, что под влиянием миграции в популяции, находящейся в работе, могут произойти значительные изменения частот проявления генов и генотипов.
Мутации
Спонтанные мутации, о которых уже упоминалось в главе 3, возникают в поколениях природных и искусственно созданных популяций с частотой около 10-6 на один ген. Принимая во внимание малую частоту мутаций, в кратковременной селекционной программе они не представляют собой фактора, требующего специального учета. Однако если идет речь о некоторой летальной или полулегальной мутации (нарушения в образовании хлорофилла и др.), вредный ген необходимо устранить с помощью отбора. Точно так же при появлении какой-то полезной мутации (устойчивость к болезни и т.п.) ее нужно отобрать для дальнейшей работы.
Отбор
Популяция состоит из различных генотипов, одни из которых живут дольше и дают больше потомства, в то время как другие живут меньше и дают малочисленное потомство. Изменение частоты проявления генов, которое происходит из-за различной продолжительности жизни и неодинаковой репродуктивной способности отдельных генотипов, в действительности и есть естественный отбор.
Генотипы с повышенной фертильностью оставляют после себя больше потомства и таким образом увеличивают частоту проявления генов, что приводит к нарушению равновесия в популяции и благоприятствует данным генотипам. Этот различный вклад генотипов следующего поколения называется адаптивной или селекционной ценностью (fitness).
Отбор может проходить на уровне фенотипа, генотипа, зиготы и гаметы, поэтому различают отбор диплоидный и гаплоидный. На каком бы уровне он ни проходил, отбор всегда приводит к изменению частоты проявления гена. Эффективность отбора зависит от уровня частоты проявления гена, на котором он протекает, от эффекта гена и интенсивности самого отбора.
Отбор на гомозиготность. Отбор по гену A, обладающему эффектом полной доминантности, охватывает генотипы АА и Аа, фенотипически не различающиеся. При наличии растений-перекрестников и случайного размножения произойдет скрещивание генотипов ААхАА, ААxАа и АаxАа и в потомстве появятся генотипы АА, АА, Аа, АА, Аа, Аа и аа. Следовательно, эффект отбора на доминантность сказывается не так быстро, как в работе с растениями-самоопылителями; в этом случае наряду с генотипами АА появляются и гетерозиготные генотипы Аа, и рецессивные генотипы аа, на которые отбор вообще не действует, но они постоянно возникают из размножаемых генотипов АахАа.
При высокой исходной частоте проявления гена А и слабой интенсивности отбора этот ген будет долго сохраняться в популяции, однако в конце концов и он элиминирует. Если при отборе какой-то ген полностью устраняется из популяции, тогда s = 1 и q = 0. Например, при завершении отбора на исключение рецессивных генов изменение частоты проявления гена составит:
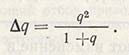
Если первоначальная частота q велика, то частота проявления гена резко упадет, в противном случае продолжительность жизни 1 + q вскоре приблизится к 1 и изменение в любом поколении вскоре станет равным q2. Поэтому при низких частотах (q = 0,01) изменение q за поколение составляет 0,0001, а это означает, что при отборе теряется только один ген на 10 тыс. Следовательно, отбор на исключение рецессивных генов при низкой их частоте в популяции - процесс чрезвычайно медленный.
Перед началом селекции растений-перекрестников часто возникает вопрос, сколько потребуется поколений для устранения из популяции какого-то рецессивного летального гена (альбинизма). При интенсивности отбора s = l (при условии, что мутации отсутствуют) для частот проявления гена в 0, 1, 2 и n поколениях отбор включает q0, q1 q2 и qn, причем:
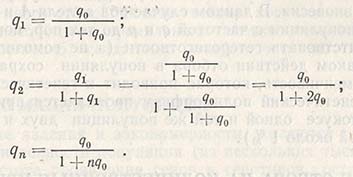
Последнюю формулу используют при расчете числа отборов в поколениях, которое необходимо для получения определенного изменения частоты проявления генов. Чтобы частота гена изменилась от 0,1 до 0,01, при s = 1 необходимо 90 поколений, а при s = 0,5 - даже 185 поколений.
Отбор на гетерозиготность. Примеры показывают, что в естественных и искусственно созданных популяциях гетерозиготы (Аа) имеют более высокую адаптивную ценность, чем гомозиготы (АА или аа). Если адаптивная ценность генотипа Аа равна единице, адаптивная ценность гомозиготы АА уменьшилась до величины коэффициента отбора, обозначенного как s, и гомозиготы аа - до величины коэффициента t, то изменение частот проявления генов, возникающее в одном поколении, рассчитывают следующим образом:
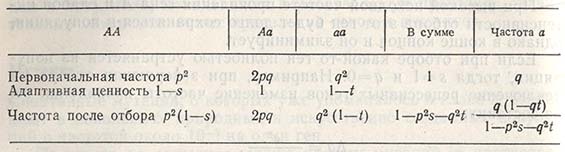
Далее из этого рассчитывают изменение в частоте проявления гена:
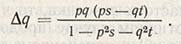
Если dq равно нулю, наступает равновесие, и частота проявления гена остается стабильной. При ps = qt частота изменяется, и величину состояния равновесия можно выразить следующим образом:
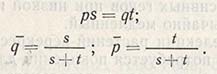
Величина состояния равновесия не зависит от первоначальной частоты гена в популяции и определяется только коэффициентом отбора не в пользу гомозигот. Следовательно, если s и t возрастают от 0 и до константных величин, р и q достигнут устойчивого состояния равновесия. В данном случае оба аллеля А и а будут сохраняться в популяции с частотой q и р до тех пор, пока отбор будет благоприятствовать гетерозиготности (а не гомозиготности).
При таком действии отбора в популяции сохраняется генетическая изменчивость, которая приводит к генетическому полиморфизму. Генетический полиморфизм проявляется при поддержании в одном локусе одной и той же популяции двух и более аллелей (с частотой около 1 %).
Влияние отбора на количественные признаки
Основное влияние отбора заключается в изменении частоты проявления генов в популяции, которая в то же время при естественном отборе всегда связана с адаптивной ценностью признака или группы признаков. Поскольку количественные признаки обусловлены большим числом генов и находятся под сильным влиянием факторов окружающей среды, действие отбора на изменение частот генов выражается через изменение фенотипической ценности как средних значений данного признака в популяции, их вариансы и ковариансы. Существуют три основные формы воздействия естественного отбора на количественные признаки в популяции: стабилизирующее, направляющее и разделяющее.
Стабилизирующий отбор. Стабилизирующий отбор (часто его называют также оптимальным, сбалансированным или отбором на среднее значение признака) наблюдается у особей со средним значением выраженности количественного признака, обладающего самой высокой адаптивной ценностью. В потомстве таких особей среднее значение признака не изменяется или же из-за утраты в ряду распределения одинаковой части крайних вариантов уменьшается его варианса.
Направляющий отбор. Вследствие отбора адаптивная ценность может возрастать или снижаться вместе со средним значением данного количественного признака. Направляющий отбор приводит к увеличению выраженности среднего значения признака в последующих поколениях, при этом варианса может измениться или же остаться постоянной.
Разделяющий отбор. Если в одной и той же популяции крайние варианты обладают лучшей адаптивной ценностью, чем варианты, имеющие обычно приближенно среднее значение адаптивной ценности, то это приводит к разделяющему отбору на плюс- и минус-варианты. При этом среднее значение адаптивной ценности может оставаться неизменным в последующих поколениях и тем не менее привести к увеличению значения вариансы.
Во всех перечисленных случаях успех отбора зависит от того, в какой мере фенотип является отражением генотипа, точнее от характера наследуемости данного количественного признака.
Отборы, проводимые в селекции растений, в большинстве своем относятся к направляющему типу, при котором особи с лучшей средней ценностью выбираются в качестве родительских форм следующего поколения. Отбор по некоторым экономически важным признакам часто бывает ограничен из-за слабой жизнеспособности чрезмерно отселектированных фенотипов как следствия стабилизирующего естественного отбора, препятствующего действию искусственного отбора. В естественных популяциях разделяющий отбор способствует проявлению генетического полиморфизма, в искусственно созданных популяциях его применяют редко.
Бесконечно малая популяция
Рассмотренные явления и закономерности касаются бесконечно большой по численности популяции (из нескольких тысяч особей), в которой частота проявления генов и генотипов остается неизменной из поколения в поколение, если на нее не воздействуют извне отбор, мутации и миграции. Эти воздействия имеют систематический и постоянный характер независимо от их направленности в сторону фиксации генов или в сторону достижения состояния равновесия. Поэтому в естественных популяциях можно предусмотреть направленность указанных воздействий.
Однако в селекции чаще всего приходится работать с небольшим числом особей, т.е. с малой (первичной, или исходной) популяцией, содержащей лишь часть генетической изменчивости. Такая популяция представляет собой лишь модель бесконечно большой популяции и характеризуется рядом отступлений и специфических особенностей в отношении тех закономерностей, которые управляют бесконечно большой популяцией.
В бесконечно малой популяции отбор уклоняющихся генотипов (точнее гамет из большой по численности популяции) ведет к изменению частоты генов из поколения в поколение, поэтому популяция эта структурно неустойчива. Фактически в ней разворачиваются дисперсные процессы, приводящие к: 1 - разделению популяции на подпопуляции, группы, линии; 2 - уменьшению генетической изменчивости и возрастанию уровня гомозиготности.
В подпопуляциях или группах все активнее идет скрещивание сходных между собой особей, т.е. инбридинг, поэтому внутри групп наблюдается уменьшение генетической изменчивости, фиксация одних и элиминация других генов и увеличение гомозиготности. Таким образом, наступает разделение между группами или линиями, которое Райт назвал случайным дрейфом (random drift).
Образование различных местных популяций у растений-самоопылителей - пример действия дисперсных процессов, происходящих в естественных популяциях. Так, по данным Бригера, в Южной Америке встречаются многочисленные местные популяции кукурузы, которые в сильной степени отличаются друг от друга, хотя при этом в пределах отдельной популяции преобладает значительная выравненность. Подобная картина характерна и для югославских местных форм ржи, особенно в горных районах: под влиянием элементов рельефа, разделяющих поля, местные популяции разбились на ряд малых популяций, внутри которых все чаще и чаще происходит скрещивание сходных между собой особей. Это привело к повышению уровня гомозиготности в популяциях и одновременно к морфологической их выравненности. Так, местная рожь из Лике фенотипически однородна, но в значительной степени отличается от местной ржи из Гламоча и в еще большей степени от местной ржи из Невесиньи.
Подобные или сходные дисперсные процессы протекают и у других видов-перекрестников: злаковых трав, бобовых и т.д. Поэтому местные популяции представляют немалый интерес для дальнейшей работы по селекции растений.
Фиксация генов
Дисперсные процессы в бесконечно малой популяции могут действовать до определенных границ, так как частота проявления гена может находиться только в пределах от 0 до 1. Так, когда ген достигает частоты 0 или 1, дальнейшее его изменение возможно лишь при мутации. Аллель, достигший частоты 1, фиксируется в популяции, достигший 0 - устраняется. В линии с фиксированным геном (принимая во внимание данный локус) все особи имеют сходный генотип. В то же время, если в линии фиксированы все локусы, наступает генетическая сбалансированность и образуется инбредная линия.
Фиксирование гена можно рассмотреть на конкретном примере. Если любая из 100 популяций начинает формироваться из двух гетерозигот (АаxАа), то потомство первого поколения будет состоять из трех генотипов: АА, Аа, аа в соотношении 1:2:1. В следующем поколении при наличии только двух родителей существует вероятность появления комбинаций из трех указанных генотипов, однако вероятность возникновения определенной комбинации зависит от частот проявления генотипов (табл. 5.2).
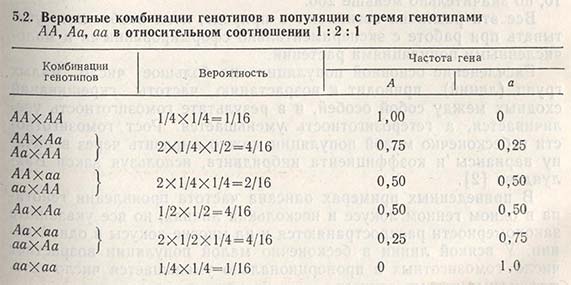
Вероятность случайного подбора обоих родителей с генотипом АА составляет 1/16, с генотипом аа - также 1/16. Это означает, что уже в первом поколении в 6 популяциях из 100 наверняка произойдет фиксация гена АА и в других 6 популяциях - фиксация гена аа. В остальных 88 популяциях, возможно, останутся следующие комбинации генотипов: ААxАа, ААxаа, AаxAа, Ааxаа. Три из этих комбинаций могут вновь дать популяции, гомозиготные по обоим аллелям, даже при условии, что в следующем поколении появится уже 15 таких популяций (15/100 = 15%). В последующих поколениях процесс будет продолжаться до тех пор, пока число популяций, достигших состояния фиксации, не станет равным 0,5N (где N - число родителей, использованных для получения поколений), т.е. при наличии двух родителей и 100 популяций 0,5N = 100. Это означает, что все 100 популяций после бесконечно большого числа поколений будут состоять из групп, достигших состояния фиксации, а частоты проявления их генов будут поделены между группой АА и группой аа пропорционально величинам р и q в основной популяции.
Таким образом, скорость фиксации гена зависит не только от частоты его проявления, но и от числа родителей в популяции. Если N равно 100 000, а первоначальная частота гена 0,5, то после продолжительного периода наступает случайная изоляция, хотя и весьма незначительная. При одинаковой частоте проявления гена и малой величине популяции от N = 1000 происходит быстрая фиксация генов и элиминация аллеля (рис. 5.2).
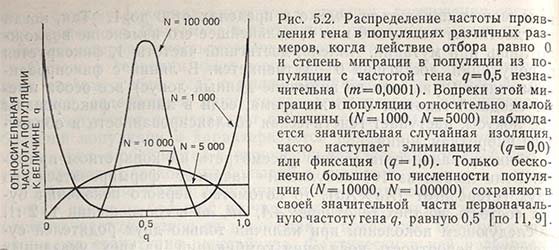
Число родителей имеет большое значение для установления генетической изоляции и выражается как эффективная величина популяции (Ne). Этот показатель отличается от действительной величины, поскольку не все члены популяции являются родителями и дают потомство. Если из 1000 особей только у 400 пар происходит скрещивание, эффективная величина популяции равна 800. При соотношении полов, не равном 1:1, наступает уменьшение эффективной величины популяции: когда на 200 особей женского пола приходится 10 особей мужского пола, Ne превышает 10, но значительно меньше 200. Все эти различия возникают в природе, и их необходимо учитывать при работе с экспериментально сформированными и малочисленными популяциями растений.
Расчленение основной популяции на большое число малых групп (линий) приводит к возрастанию частоты скрещиваний сходных между собой особей, и в результате гомозиготность увеличивается, а гетерозиготность уменьшается. Рост гомозиготности в бесконечно малой популяции можно выразить через величину вариансы и коэффициента инбридинга, используя закон Вахлундова.
В приведенных примерах описана частота проявления генотипа в одном генном локусе и нескольких линиях, но все указанные закономерности распространяются и на многие локусы в одной линии. У всякой линии в бесконечно малой популяции возрастает число гомозиготных и пропорционально уменьшается число гетерозиготных генных локусов.
Рассмотренный в главе общий характер дисперсных процессов показывает, что дифференциация популяций на уровне подлиний вследствие случайной изоляции и фиксации генов приводит к сокращению масштабов изменчивости, росту гомозиготности и сбалансированности подлиний. Полной гомозиготности бесконечно малая популяция растения-перекрестника достигает в процессе инбридинга, если он возможен у данного вида.