Вертикальная и горизонтальная устойчивость
Ван дер Планк называет вертикальной устойчивостью тот случай, когда отдельный сорт устойчив к одним расам патогена и неустойчив к другим расам того же самого патогена. Если сорт одинаково устойчив ко всем расам, Ван дер Планк говорит о горизонтальной устойчивости.
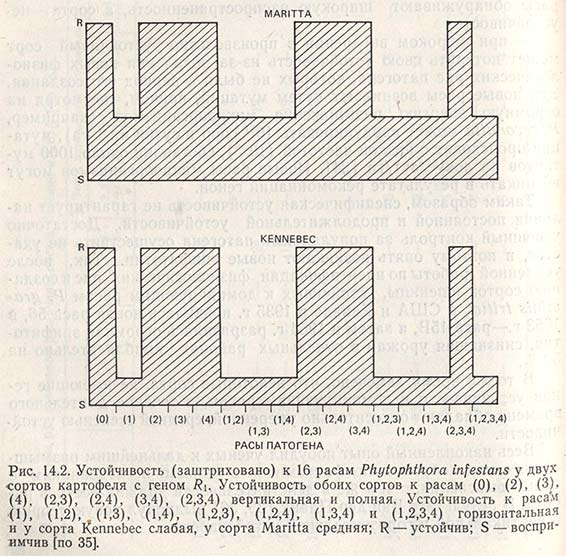
На примере устойчивости сортов картофеля к фитофторозу (возб. Phytophthora infestans) Ван дер Планк показывает, чем различаются между собой вертикальная и горизонтальная устойчивости (рис. 14.2). Сорта картофеля Kennebec и Mariita обладают геном R1 который определяет их вертикальную устойчивость к расам, (0), (2), (3), (4), (2,3), (2,4), (3,4) и (2,3,4). Однако в реакции на остальные расы (1), (1, 2) и т.д., против которых ген R1 не дает устойчивости, между сортами существуют различия. Инфекция быстро развивается на сорте Kennebec, и когда листья растений этого сорта уже становятся бурыми, у растений сорта Maritta они еще сохраняют зеленый цвет. Можно с уверенностью сказать, что Maritta обладает более высокой, чем Kennebes, горизонтальной устойчивостью (которую называют также полевой устойчивостью) (см. рис. 14.2). у сортов, обладающих другими генами устойчивости (R2, R3, R4) или их комбинациями, картина в целом такая же, как и у сортов с генами R1.
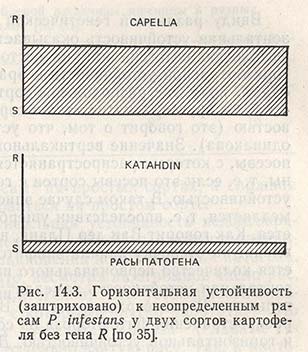
Во всяком случае, некоторые сорта не имеют генов устойчивости R и, следовательно, не обладают вертикальной устойчивостью, но это не означает, что все они восприимчив вы к патогену и между ними нет различий. Сорт картофеля Kathadin очень восприимчив к фитофторозу листьев и в полевых условиях обнаруживает низкую горизонтальную устойчивость, в то время как Capella намного успешнее сопротивляется инвазии. У этого сорта пораженность, проявляющаяся в виде некротических пятен на листьях, растет очень слабо, листья и стебли остаются долго зелеными даже в благоприятных для развития фитофтороза полевых условиях. Можно сказать, что сорт Capella имеет высокую горизонтальную устойчивость (рис. 14.3).
Используя эти и другие примеры, Ван дер Планк дает объяснение генетических основ вертикальной и горизонтальной устойчивости. Устойчивость, обусловленная одним геном (моногены) или несколькими генами (олигогены) с сильно выраженным эффектом действия (так называемые главные гены), относится к вертикальной. Если у патогена отсутствуют гены вирулентности, устойчивость обнаруживается. Если же патоген имеет гены вирулентности, а растения не несут генов устойчивости, возникает заболевание. В этом и заключается взаимоотношение по принципу ген-на-ген, а отсюда берет свое начало специфическая устойчивость к отдельным расам. Таким образом, можно предположить, что выражение вертикальная устойчивость есть синоним специфической устойчивости.
Наоборот, устойчивость, обусловленная большим числом генов (полигены), обладающих каждый в отдельности малым эффектом действия (так называемые малые гены), но в целом оказывающих кумулятивное действие, называется горизонтальной. Такая устойчивость, по мнению Ван дер Планка, не зависит от системы ген-на-ген и поэтому является неспецифической (race-non-specific).
Между этими двумя типами устойчивости не существует резкого разграничения. Одни сорта обладают вертикальной и горизонтальной устойчивостью, другие обладают только одним из указанных типов устойчивости.
Ввиду различной генетической природы вертикальная и горизонтальная устойчивость оказывает различное воздействие на эпифитологию патогена или на состояние посевов в поле. Значение горизонтальной устойчивости возрастает, если патоген переходит с одного поля, где расположены сорта с горизонтальной устойчивостью, на другое, занятое также сортами с горизонтальной устойчивостью (это говорит о том, что устойчивость сортов генетически одинакова). Значение вертикальной устойчивости возрастает, если посевы, с которых распространяется инфекция, генетически различны, т.е. если это посевы сортов с генетически разной вертикальной устойчивостью. В таком случае эпифитотия останавливается или замедляется, т.е. впоследствии ущерб от действия патогена уменьшается. Как говорит Ван дер Планк, вертикальная устойчивость сорта влияет на отсрочку начала эпифитотии, и таким образом уменьшается количество первоначального инокулюма, в то время как горизонтальная устойчивость замедляет рост эпифитотии уже после ее вспышки. В подтверждение этого Ван дер Планк приводит пример распространения фитофтороза на сортах картофеля с вертикальной и горизонтальной устойчивостью. Для того чтобы степень заражения шести сортов с генами R повысилась от 1 до 3 баллов, в среднем потребовалось 15,5 дня, а на шести сортах, не имевших генов R, этот период составил 26,7 дня.
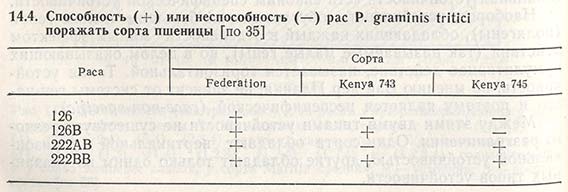
В связи с сильным давлением отбора, способствующим проявлению вертикальной устойчивости, интересно подчеркнуть, что она сохраняется наиболее долго при наличии простых рас (с одним или парой генов) и в простых сортах (с одним, парой генов или без генов устойчивости). Исследования Уотсона и Сингха показали, что самая простая раса 126 P. graminis tritici могла вызвать поражение сорта пшеницы Federation, но не сортов Kenya 743 и 745, а сама сложная раса 222ВВ не поражала все три сорта (табл. 14.4). Вначале раса 126 была самой распространенной, но когда в производство поступили генетически различающиеся сорта, характер распространенности расы изменился. Так, в районе Гуннедах, где возделывали сорта, устойчивые к расам 126 и 126В, новое направление отбора привело к тому, что доминирующими стали расы 222АВ и 222ВВ (табл. 14.5). Между тем в районах Вагга и Темора, где в основном выращивали неустойчивые сорта, раса 126 так и осталась доминирующей. Следовательно, стабилизирующий отбор благоприятствовал простым расам на простых сортах, а сложная раса 222ВВ не смогла адаптироваться на простом сорте.

Как уже указывалось, вертикальная устойчивость недолговременна из-за постоянного изменения в характере распространенности физиологических рас и возникновения новых мутаций патогена. К этому еще нужно добавить, что, несмотря на огромное число рас, идентифицированных у многих патогенов, гены устойчивости главным образом входят в те расы, с которыми «легко работать». Это гены, эффект действия которых бросается в глаза (главные гены), - один или пара генов на отдельно взятую расу. Так, к настоящему времени установлено, что устойчивость к P. graminis triiici определяется всего двумя десятками генов (гены Sr), а число идентифицированных рас этого патогена достигает 300. Устойчивость к большинству этих рас обусловлена, вероятно, большим числом генов (полигены), из которых ни один не обладает таким эффектом выраженности, чтобы его можно было бы идентифицировать в отдельности. Такова в действительности устойчивость к большинству рас, оставшаяся от естественного отбора, и селекционеры, отбирающие растения с низкой степенью зараженности, не допускают или замедляют развитие патогена с помощью так называемой полевой устойчивости, являющейся синонимом горизонтальной устойчивости. Поэтому, хотя работа над созданием горизонтальной устойчивости не носила планового характера, определенное число сортов, широко используемых в производстве, обладает этим типом устойчивости, а некоторые из них имеют и горизонтальную, и вертикальную устойчивость.
Теория Ван дер Планка о вертикальной и горизонтальной устойчивости имела чрезвычайно большое значение для всестороннего и глубокого осмысления сущности устойчивости растений к патогенам и в течение последнего десятилетия оказала сильное влияние на весь ход селекционной работы, связанной с устойчивостью к болезням. Исследования специфической устойчивости в значительной мере уступили место исследованиям неспецифической, или горизонтальной, устойчивости. Особенно большой вклад в дальнейшую разработку теории Ван дер Планка внес Робинсон, который дал объяснение многим понятиям, а также предложил новые термины и дал примеры. Генетическое объяснение устойчивости еще не нашло общего признания, и поэтому оно непрерывно вызывает споры.
Парлевлиет и Цадокс полагают, что разделение устойчивости на горизонтальную (которой приписывают постоянство, полигенный характер наследования и расовую неспецифичность) и вертикальную (для которой характерны непостоянство, моногенное наследование и расовая специфичность) приводит к идее о существовании не только двух типов устойчивости, но и двух типов генов устойчивости. Возможно, такая классификация удобна при проведении исследований, но трудно представить себе, что она существует в природе.
Более того, допустив, что большинство генов устойчивости - это главные гены, функционирующие на основе системы ген-на-ген, нет причины отрицать, что и малые гены хозяина взаимодействуют по этому же принципу с малыми генами патогена. Если эффект действия главных генов хозяина по отношению к расам патогена относительно легко подвергнуть проверке, то гены, имеющие аддитивный эффект, экспериментально испытывать по принципу ген-на-ген весьма трудно. Поэтому Парлевлиет и Цадокс разработали модель для испытания генов с малым эффектом. При этом предусматривается два варианта.
1) Аддитивная модель. У растений-хозяев и их патогенов, а также между растениями-хозяевами и между патогенами плюс-аллели имеют аддитивный эффект. Каждый плюс-аллель хозяина повышает его устойчивость на 10%, а каждый плюс-аллель патогена снижает устойчивость хозяина на 10 %. несмотря на локус, в котором находится плюс-аллель. Очевидно, гены хозяина и патогена взаимодействуют не по системе ген-на-ген, а неспецифично, что, по Ван дер Планку, характеризует горизонтальный тип устойчивости.
2) Модель взаимодействия. Эта модель также допускает аддитивный эффект генов, но гены хозяина взаимодействуют с генами патогена на основе системы ген-на-ген. Один плюс-аллель в локусе хозяина с минус-аллелем соответствующего локуса патогена увеличивает устойчивость на 10%. Если хозяин имеет только минус-аллель, устойчивость будет отсутствовать даже при условии, что патоген в соответствующем локусе имеет минус- или плюс-аллель.
В таблице 14.6 приведено несколько комбинаций взаимоотношения хозяин - патоген. Согласно аддитивной модели, степень заражения не зависит от идентичности локуса, в котором находятся плюс-аллели патогенности (последние три комбинации). Согласно модели взаимодействия, или модели ген-на-ген, степень заражения зависит от идентичности локуса, в котором находятся плюс-аллели патогенности (последние три комбинации).
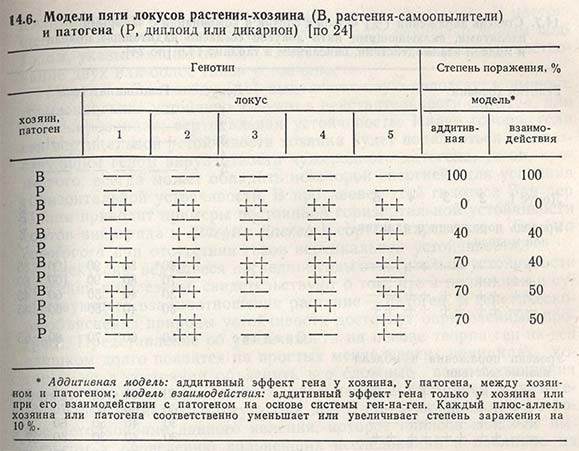
Для более четкого различия вертикальной и горизонтальной устойчивости Парлевлиет и Цадокс приводят также реакцию четырех сортов на четыре изолята, учитывая случай аддитивной и взаимодействующей модели (табл. 14.7). Полученные данные показывают, что вертикальную устойчивость нельзя отличить от горизонтальной, если гены устойчивости имеют аддитивный эффект. В модели взаимодействия степень заражения одного сорта зависит не только от числа плюс-генов устойчивости и плюс-генов патогенности, но также и от идентичности локусов, содержащих плюс-гены.

Что касается мутаций от минус- к плюс-аллелям патогенности, то, согласно модели взаимодействия, они достигнут степени выраженности только тогда, когда соответствующий локус хозяина по плюс-аллелям будет нейтрализован. Это значит, что модель, основанная на специфичных малых генах, более стабильна, чем модель, базирующаяся на неспецифичных малых генах.
По мнению Парлевлиета и Цадокса, связь хозяин - патоген, основанная на системе ген-на-ген, может в природе привести к устойчивому равновесию, полезному как для хозяина, так и для патогена. Если находящаяся в равновесии популяция патогена дает возможность хозяину поддерживать устойчивость к болезни, то сам патоген не в состоянии выжить при гибели хозяина. Приспособительная сила популяции патогена пропорциональна силе устойчивости популяции хозяина, и наоборот. Устойчивость популяции хозяина - это результат кумулятивного эффекта всех генов устойчивости в популяции.
В конце концов, Парлевлиет и Цадокс пришли к заключению, что горизонтальная и вертикальная устойчивость, как предполагал Ван дер Планк, представляет собой не два различных типа устойчивости, а только одну полигенную, точнее олигогенную, устойчивость. У этой устойчивости единичные гены хозяина взаимодействуют специфическим образом на основе системы ген-на-ген с генами вирулентности патогена.
В своем дальнейшем разъяснении генетических основ вертикальной и горизонтальной устойчивости Ван дер Планк не согласился с гипотезой Парлевлиета и Цадокса о том, что горизонтальная устойчивость - это кумулятивная вертикальная устойчивость. Моногенность вертикальной устойчивости и полигенность горизонтальной не объясняют различий между ними, поскольку горизонтальная устойчивость также может быть моногенной, и если вертикальную устойчивость можно объяснить с позиций принципа ген-на-ген, то горизонтальную таким образом интерпретировать нельзя. Сущность состоит в различиях, возникающих при кодировании первичных белков хозяина и патогена на молекулярном уровне. Ван дер Планк выдвигает гипотезу, в соответствии с которой вертикальную устойчивость, точнее восприимчивость, определяет процесс полимеризации, а горизонтальную устойчивость, или восприимчивость, - процесс катализа или его продукты. В настоящее время такое представление является совершенно новым подходом, указывающим на то, что нет оснований отвергать существование двух или более типов устойчивости.
Мартин и Эллингбоу выдвигают иную гипотезу, а именно: горизонтальная устойчивость - это в действительности скрытая, или замаскированная, вертикальная устойчивость. Иначе говоря, если ген вертикальной устойчивости хозяина будет подавляться соответствующим геном вирулентности чужеродного патогена, то он, кроме того, всегда может обладать некоторой энергией для усиления горизонтальной устойчивости. В противовес этой гипотезе Ван дер Планк приводит примеры постоянной горизонтальной устойчивости сортов винограда к Boirytis cinerea и сортов картофеля к Erwinia carotovora при отсутствии генов вертикальной устойчивости.
Дискуссии, ведущиеся последние годы относительно устойчивости растений к болезням, свидетельствуют о том, что в признании и существующего взаимоотношения растение - патоген, и генетического объяснения природы устойчивости достигнут определенный прогресс. Представление об устойчивости на основе теории ген-на-ген слишком долго покоится на простых менделевских принципах, которые не в состоянии объяснить все сложные взаимоотношения между двумя видами живых организмов - растением и патогеном. Поэтому велика заслуга Ван дер Планка, предложившего новое истолкование данного явления, которое явилось мощным импульсом к проведению дальнейших исследований и повлияло на переориентацию селекции растений, занимающейся вопросами устойчивости к паразитическим организмам.