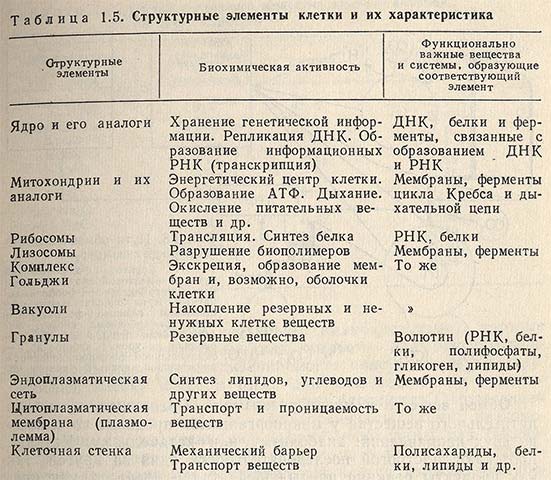
Питание микроорганизмов
Непропорционально большие количества различных веществ, перерабатываемых ничтожно малыми по величине микроорганизмами, позволили еще Пастеру говорить о «бесконечно большой роли бесконечно малых существ». Бактериальная клетка может потреблять за сутки количество пищи, превышающее в 30-40 раз ее собственную массу. Поступление питательных веществ и выделение продуктов жизнедеятельности осуществляются с большой скоростью всей поверхностью (диффузия) микробной клетки. Происходит интенсивный обмен между клеткой и внешней средой. Этот обмен состоит из двух процессов: получение необходимых для роста питательных веществ из окружающей среды, синтез из них структурных частей клетки (питание) и выделение в окружающую среду конечных продуктов метаболизма. В табл. 1.5 приведены состав микробной клетки, а также вещества и системы, ответственные за образование структурных элементов.
Возможность использования микроорганизмами для питания различных химических соединений определяется двумя основными факторами. Первый из них - способность вещества проникать внутрь клетки через цитоплазматическую мембрану. Механизм, с помощью которого осуществляется перенос растворенных веществ через мембрану, отличается высокой избирательностью.
Молекула растворенного вещества может проникать через липопротеидную мембрану путем пассивной диффузии или как большинство растворенных веществ с помощью специфических механизмов переноса. Считают, что макромолекулы-переносчики расположены в мембране. Они захватывают молекулу растворенного вещества и переносят ее к внутренней поверхности мембраны, где высвобождают ее в цитоплазму. Такие связанные с мембраной молекулы-переносчики называются пермеазами. В настоящее время считают, что пермеазы - это вещества ферментной природы.
Многие микроорганизмы непроницаемы для органических кислот трикарбонового цикла, в то время как сахара обычно проникают в клетку достаточно легко. Важным фактором, влияющим на способность молекулы проникать внутрь клетки, является ее размер. Для многих высокомолекулярных соединений цитоплазматическая мембрана оказывается непроницаемой. Однако это не означает, что все высокомолекулярные соединения не могут быть использованы микроорганизмами в качестве питательных веществ.
Многие микроорганизмы выделяют различные экзогенные ферменты, вызывающие гидролиз молекул, размеры которых слишком велики для проникновения через цитоплазматическую мембрану. Наиболее широко распространены ферменты, катализирующие гидролиз таких полимеров, как полисахариды и белки, а также молекул, имеющих тенденцию к агрегации в крупные мицеллы (липиды, фосфолипиды). Не все гидролитические ферменты, выделяемые микроорганизмами, диффундируют в среду. Некоторые из них остаются тесно связанными с микробной клеткой, находясь либо внутри клеточной стенки, либо на внешней стороне цитоплазматической мембраны. Химические соединения, которые способны проникать внутрь микроорганизма, могут быть использованы только как питательные вещества, когда этот микроорганизм синтезирует ферменты, необходимые для их метаболизма.
Обмен веществ у микроорганизмов
Превращение любого питательного вещества у микроорганизмов проходит по одному из двух направлений: анаболизму или катаболизму. Реакции протекают в строгой последовательности одна за другой, так как продукты реакции предыдущей стадии процесса, как правило, служат субстратом для последующей.
На рис. 1.8 представлен путь обмена веществ у микроорганизмов. Он состоит из реакций, которые поставляют клеткам строительный материал, и реакций, которые обеспечивают все превращения в клетке, т. е. реакций, ответственных за конструктивный и энергетический обмены.
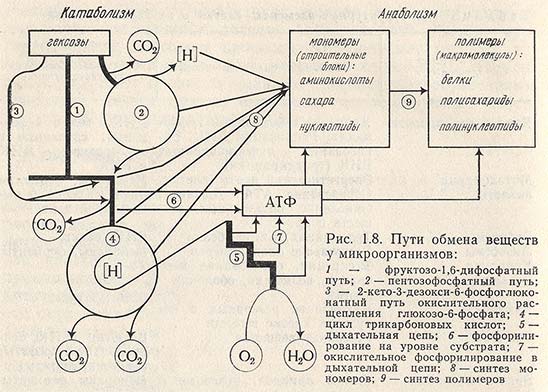
При анаболических превращениях обычно наблюдают синтез новых клеточных компонентов, тогда как результатом катаболизма является образование низкомолекулярных соединений. Одни из этих соединений - некоторые продукты обмена - выделяются из клетки, другие служат предшественниками для биосинтетических реакций и, таким образом, включаются в анаболические пути. Катаболические превращения часто служат также источником необходимой для метаболизма клетки энергии в форме АТФ или некоторых других богатых энергией соединений, которые могут быть использованы в реакциях биосинтеза. Все это еще раз свидетельствует о существующей тесной связи между анаболическими и катаболическими путями метаболизма.
Основным условием для биосинтеза клеточных компонентов является снабжение клетки соответствующими низкомолекулярными соединениями (сахарами, аминокислотами), которые служат предшественниками, или «сырьем» для биосинтетических процессов. Эти соединения могут присутствовать в окружающей клетку среде и тогда они непосредственно вовлекаются организмом в те или иные пути биосинтеза. Окружающая среда не может снабдить микроорганизмы всеми низкомолекулярными соединениями, которые необходимы для биосинтетических реакций, поэтому некоторые, а часто и все подобного рода соединения бактериальная клетка должна «выработать» из питательных веществ, имеющихся в ее распоряжении. В процессе роста наряду с клеточными компонентами микроорганизм образует в незначительных количествах вещества сложной молекулярной структуры - различные ферменты, антибиотики, витамины и другие биологически активные вещества.
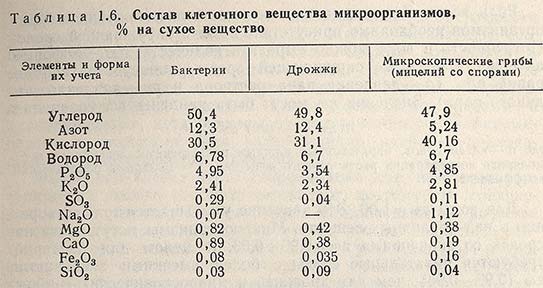
Микроорганизмам для формирования новой клетки необходимо поступление из окружающей среды различных питательных компонентов в доступной форме в определенных качественных и количественных соотношениях. Все элементы, входящие в состав клеточного вещества (табл. 1.6), должны находиться в питательной среде.
Без наличия хотя бы одного из указанных элементов рост будет незначительным или его совсем не будет. Например, если из полноценной питательной среды полностью изъять цинк, то Asp. niger образует только 0,5 мг сухой массы мицелия. При наличии цинка грибная пленка достигает нескольких граммов. Синтез клеточного вещества ускоряется во много раз потому, что цинк, как и многие другие микроэлементы, входит в состав ферментов и без него становится невозможной нормальная физиологическая активность клетки.
Роль воды. Для нормального роста и размножения микроорганизмов необходимо присутствие воды в окружающей среде. Потребность в воде можно выразить количественно - в форме активности воды aw окружающей среды или субстрата; она равна р/р0 (р - давление пара раствора и р0 - давление водяного пара). Значения aw могут быть найдены из уравнения
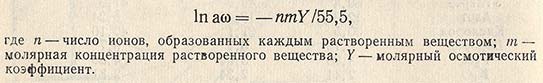
Для воды aw = 1,00. Это значение уменьшается при растворении в ней различных веществ. Микроорганизмы могут расти на средах со значениями aw = 0,99 / 0,63. В целом для бактерий требуются питательные среды с более высокими значениями aw (0,99-0,93), чем для дрожжей и микроскопических грибов. Значения aw, оптимальные для роста дрожжей, варьируют, но минимальные величины для этих организмов (0,91-0,88) ниже, чем для большинства бактерий. Микроскопические грибы, в общем, более способны переносить высушивание, чем другие микроорганизмы, и для некоторых штаммов, например для представителей группы Asp. glaucus, нижний предел значения aw около 0,60.
Источники углерода. Среди всех элементов, используемых микроорганизмами в качестве питательных веществ, наиболее важное значение имеет углерод, содержание которого в пересчете на сухую массу микроорганизмов составляет примерно 50% (см. табл. 1.6). Микроорганизмы могут использовать самое простое углеродное соединение - диоксид углерода. Для некоторых из них С02 является единственным источником углерода. Такие микроорганизмы носят название автотрофов. Микроорганизмы, которым помимо диоксида углерода необходимо присутствие в среде некоторых органических источников углерода, называются гетеротрофами.
Количество органических соединений, используемых гетеротрофными микроорганизмами в качестве источников углерода, чрезвычайно велико. Наиболее доступными являются полуокисленные атомы углерода: -СН2ОН, -СНОН-, =СОН-. Поэтому различные сахара, глицерин, маннит, органические кислоты характеризуются высокой питательной ценностью. Плохо усваиваются микроорганизмами более окисленные (группа -СООН) соединения. Какое бы углеродное соединение ни использовалось микробной клеткой, оно сначала расщепляется до низкомолекулярных веществ, которые затем вовлекаются в тот или иной биосинтетический процесс. Во время процесса расщепления из химических соединений извлекается энергия, которая в форме АТФ может использоваться микроорганизмами.
К наиболее доступным для микроорганизмов источникам углерода относятся углеводы, прежде всего моносахариды, особенно гексозы. Среди пентоз наиболее широко потребляемым сахаром, по-видимому, является ксилоза. Источником углерода часто могут служить многоатомные спирты. Маннит, например, легко потребляется многими микроскопическими грибами и актиномицетами, хотя в целом он плохо используется фикомицетами и дрожжами. Для актиномицетов наиболее подходящий источник углерода - глицерин.
Микробная клетка часто непроницаема для некоторых органических кислот, особенно кетокислот, поэтому эти соединения в ряде случаев не могут быть использованы в качестве источников углерода. Следует отметить, что использование нейтральных органических кислот вызывает увеличение pH в процессе роста, что также неблагоприятно сказывается на росте культуры. Некоторые органические кислоты (например, лимонная и винная) являются хелатными агентами, и, таким образом, недостаток ионов металла может влиять на рост микроорганизмов.
Известны некоторые микроорганизмы, например представители рода Pseudomonas, для которых источником углерода служат липиды. В результате гидролиза липидов ферментом липазой образуются глицерин и жирные кислоты. Оба эти компонента или один из них могут быть источником углерода.
Некоторые микроорганизмы (род Candida, Torulopsis, Rhodotorula и др.), объединяемые в отдельную физиологическую группу - углеводородные микроорганизмы, способны окислять углеводороды. Последовательность этапов окисления представлена на рис. 1.9.
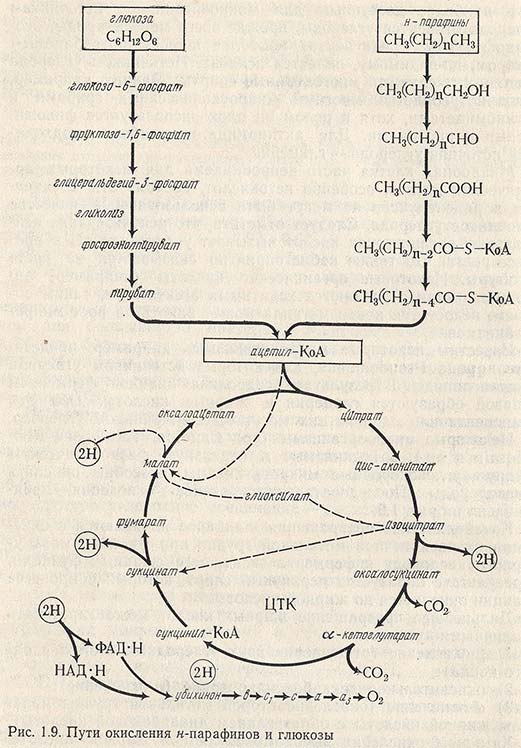
Катаболические превращения н-алканов начинаются с окисления их по конечной метальной группе при участии молекулярного кислорода и фермента оксидазы смешанных функций, в результате образуется первичный спирт, который в процессе реакции окисляется до жирной кислоты.
Дальнейшее превращение жирных кислот может протекать различными путями:
1) B-окисление (отщепление двух углеродных атомов с одного конца);
2) окислительное декарбоксилирование (а-окисление);
3) w-окисление (окисление второй метильной группы молекулы жирной кислоты с образованием дикарбоновой кислоты).
Химизм B-окисления высокомолекулярной жирной кислоты заключается в последовательном отщеплении двух углеродных атомов с образованием уксусной кислоты и новой жирной кислоты, которая подвергается B-окислению.
Окисление жирной кислоты возможно только после ее активации путем этерификации с образованием соответствующих КоА-эфиров в присутствии АТФ.
B-окисление - последовательное окислительное отщепление молекул ацетил-КоА от КоА-эфиров насыщенных жирных кислот - осуществляется в митохондриальном матриксе.
Согласно современным представлениям, отщепление каждой молекулы ацетил-КоА включает четыре стадии:
1) дегидрирование по атомам углерода в положениях 2 и 3 катализируемое ФАД-зависимой ацил-КоА-дегидрогеназой;
2) гидратация образующейся на первой стадии 2,3-транс-двойной связи, катализируемая эноилгидратазой;
3) дегидрирование образующегося на второй стадии КоА-эфира L-B-оксикислоты, катализируемое НАД-зависимой дегидрогеназой;
4) расщепление (тиолиз) образовавшегося на третьей стадии КоА-эфира B-кетокислоты, протекающее в присутствии КоА и приводящее к образованию ацетил-КоА и КоА-эфира жирной кислоты, укороченной по сравнению с исходной на два атома углерода.
Далее укороченный КоА-эфир жирной кислоты становится исходным соединением для аналогичной последовательности реакций, в результате которой образуется еще одна молекула ацетил-КоА-эфира жирной кислоты, укороченная еще на два атома углерода. Этот цикл реакций повторяется до полного окисления исходной жирной кислоты.
Таким образом, в результате B-окисления жирной кислоты, полученной при первичном окислении н-алкана, образуется ацетил-КоА, который далее подвергается окислению до СO2 и Н2O согласно циклу трикарбоновых кислот.
Источники азота. Для развития микроорганизмов, а следовательно, синтеза белковых веществ и построения протоплазмы живой клетки необходимы различные источники азота.
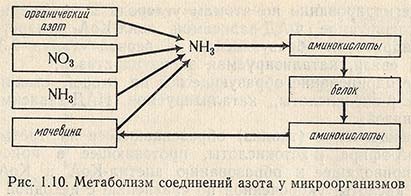
На рис. 1.10 представлены пути метаболизма соединений азота у микроорганизмов, в частности у микроскопических грибов. Наиболее доступными из них являются ионы аммония NH4+ и аммиак NH3. Они проникают в бактериальную клетку, где сравнительно легко преобразуются в имино-(-NH-) и аминогруппы (-NH2) в молекулах аминокислот, гетероциклических оснований и других химических соединений, входящих в состав протоплазмы. Наиболее универсальным источником азота для микроорганизмов является аммоний. Как источник азотного питания свободный аммиак обычно непригоден для микроорганизмов, так как при его накоплении наступает отравление клетки. Микроорганизмы используют аммонийные соли, например (NH4)2SO4, NH4Cl, которые по мере потребления иона аммония при отсутствии нейтрализующего агента подкисляют среду, что не способствует нормальному развитию клеток. Микроскопические грибы, актиномицеты, водоросли усваивают нитраты. Азот нитратов должен быть предварительно восстановлен до аммиака и только после этого он ассимилируется микроорганизмами. Нитраты необходимы тем микроорганизмам, которые боятся подкисления, так как при использовании NO3- остаются ионы металлов Na+ и К+, которые создают щелочные условия среды. Микроорганизмы в качестве источника азота могут также использовать белки, аминокислоты, пептон, пептиды. Белки усваивают только те микроорганизмы, которые выделяют в среду ферменты, расщепляющие белковую молекулу до пептидов и аминокислот. Некоторые аминокислоты необходимы самим микроорганизмам, другие - являются стимуляторами роста, присутствие третьих вовсе необязательно: микробная клетка синтезирует их сама из безазотистых органических веществ и аммиака. Например, для роста микроскопических грибов наилучшими источниками азота являются кислоты аспарагиновая, глутаминовая и их амиды, глицин; менее пригодны серосодержащие аминокислоты (цистин, цистеин, метионин) и циклические (триптофан). Особым случаем азотного питания является использование некоторыми видами синезеленых водорослей, бактериями, актиномицетами, микроскопическими грибами газообразного азота, состояние окисленности которого равно 0.
Источники фосфора. Фосфор необходим всем живым клеткам микроорганизмов. Фосфор входит в состав наиболее важных соединений протоплазмы (фосфолипидов, коферментов, нуклеиновых кислот), определяющих ее структуру и функции. В отличие от других элементов (азота и серы) фосфор входит в состав органического вещества клетки только в окисленном (в форме Р2О5) состоянии. Фосфор не вступает в соединения с углеродом, а образует с ним связи (при большой затрате энергии в АТФ) только через атомы азота или кислорода. В связи с этим некоторые органические соединения фосфора используются микроорганизмами как аккумуляторы энергии, освобождающейся при окислительных процессах. Для нормального роста и жизнедеятельности микробной клетки в окружающей среде должны присутствовать фосфаты, обычно неорганические одно- или двузамещенные фосфаты калия или натрия. В некоторых питательных средах источником фосфатов могут служить нуклеиновые кислоты.
Источники серы. Они играют большую роль в структуре клеток, так как входят в состав белков в виде серосодержащих аминокислот: цистина, цистеина, метионина. Для большинства микроорганизмов хорошим источником серы служат сернокислые соли которые в процессе ассимиляции восстанавливаются И используются в синтезе аминокислот, содержащих серу.
Аминокислота цистеин, которая присутствует в форме аминокислотного остатка в белках, является наиболее важным серосодержащим компонентом клетки. Атом серы в цистеине входит в состав тиоловой (-SH) группы. Атомы серы в большинстве других серосодержащих компонентов клетки (метионин, биотин, тиамин, глутатион) образуются из -SH-группы цистеина. Сера входит в состав биотина и тиамина, а также коэнзима А, участвующего в обмене жирных кислот и липидов в цикле Кребса. Другая функция серы состоит также в стимуляции протеолитических ферментов. Возможно, что воздействие на эти ферменты состоит в участии содержащихся -SH-групп сульфгидрильных соединений в процессах деления клеток. Этих соединений обычно бывает очень много в молодых, растущих клетках. Большинство микроорганизмов используют серу из окружающей среды в форме сульфат-иона, в котором атом серы находится в окисленном состоянии (+6) и должен быть восстановлен клеткой до состояния -2 (как в группе -SH). Для многих микроорганизмов единственным источником серы может служить тиосульфат-ион (S2O3-2). Некоторые микроорганизмы, утратившие способность восстанавливать сульфаты, нуждаются в соединениях, содержащих атом серы в восстановленной форме, как, например, сероводород, цистеин.
Факторы роста
Микроорганизмы, лишенные способности синтезировать некоторые органические соединения (аминокислоты, пуриновые и пиримидиновые основания, витамины), необходимые для построения нового клеточного материала, называются ауксотрофами. Ауксотрофные свойства микроорганизмов связаны с нарушением нормальных биосинтетических процессов. Последние в свою очередь связаны с блокированием специфических химических реакций превращения веществ вследствие нарушения активности соответствующих ферментных систем. Для нормального роста и жизнедеятельности ауксотрофов соединения (факторы роста), синтез которых нарушен, необходимо вводить в питательные среды. Следует отметить, что одно и то же вещество для различных микроорганизмов может быть необязательным, или действовать как стимулятор, или быть необходимой составной частью питательной среды. Факторы роста микроорганизмов нельзя рассматривать, как катализаторы процессов роста. Эти соединения необходимы клеткам в малых дозах и используются для синтеза физиологически активных веществ, регулирующих внутриклеточный метаболизм. Потребность микроорганизмов в факторах роста не является постоянной, она может меняться в зависимости от условий их культивирования.
Микроорганизмы могут быть ауксотрофными по одной или более, из 20 аминокислот, которые входят в состав белков (рис. 1.11). Обычно микроорганизмы нуждаются в L-аминокислотах, но для некоторых из них характерна потребность в D-аланине, необходимом, вероятно, для синтеза гликопептида клеточной стенки. Концентрации аминокислот, обеспечивающие максимальный рост ауксотрофов, обычно находятся в пределах 20-50 мкг на 1 мл.
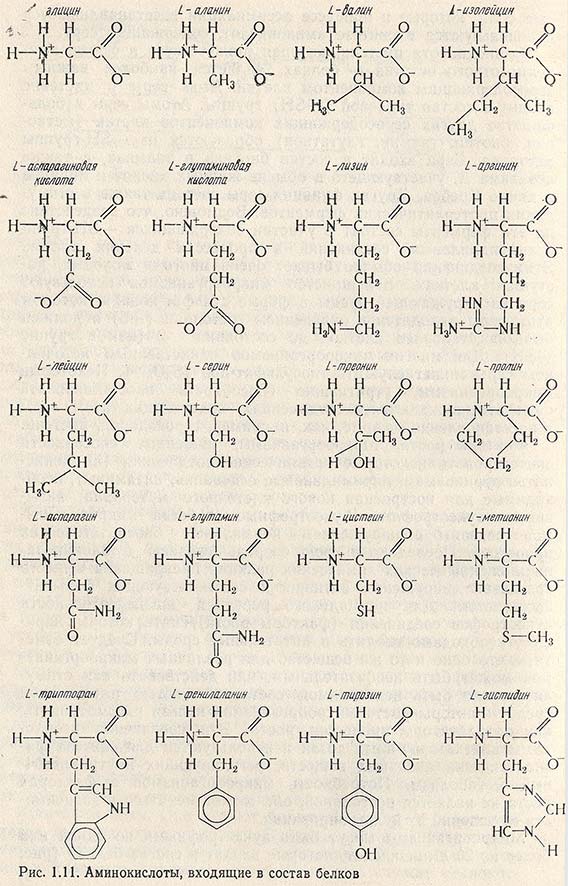
Некоторые микроорганизмы нуждаются в пуриновых и пиримидиновых основаниях или их производных, входящих в состав нуклеиновых кислот. Эти вещества нужны в основном в качестве строительного материала для нуклеотидов. Одно из пуриновых оснований - аденин - входит не только в нуклеиновые кислоты, но и в состав коэнзимов, в частности коэнзима А (КоА). Концентрации пуринов и пиримидинов, необходимые для максимального роста микроорганизмов, обычно составляют 10-20 мкг/мл.
Витамины группы В были первыми факторами роста, потребность в которых была обнаружена у микроорганизмов. Каждый из витаминов выполняет определенную биохимическую функцию в микробной клетке. Почти все витамины группы В являются активными группами ферментов (играют роль коферментов или входят в их состав), участвующих в переносе атомов водорода, остатка фосфорной кислоты или групп CH3 и т. д. Отдельные витамины не только выполняют в жизни микроорганизмов биохимические функции, но и используются для синтеза нуклеопротеидов. В связи с этим отсутствие витаминов в микробной клетке приводит к нарушениям обмена веществ и делает рост микроорганизмов невозможным. Известно около 15 витаминов, по которым микроорганизмы могут быть ауксотрофными. Концентрации витаминов, необходимые для максимального роста микроорганизмов, различны, но довольно низки (обычно 1-50 мкг/мл).
Для синтеза клеточных компонентов необходимы различные вещества. В питательной среде должны присутствовать все элементы, из которых строится клетка, и притом в таких количествах и форме, которые микроорганизмы способны усваивать. Количество минеральных веществ, содержащихся в бактериальных клетках, составляет 2-14 % к их сухой массе. В больших количествах (около 10-3 – 10-4 М) необходимы углерод, водород, кислород, азот, фосфор, сера, калий, кальций, железо, магний.
Многим микроорганизмам необходимы также и микроэлементы (около 10-6 – 10-8 М): медь, цинк, кобальт, никель, хлор, натрий, кремний, молибден, марганец и др. Элементы минерального питания выполняют не только структурную, но и функциональную роль. Железо, например, необходимо для синтеза пероксидазы, каталазы, тогда как цинк способствует превращению продуктов расщепления углеводов в составные части клетки. Магний активирует целую группу ферментов, ответственных за процессы брожения и дыхания. Кальций может влиять на избирательную сорбцию клеток, снижая проницаемость протоплазмы.