Кинетика роста микроорганизмов
Основной стадией любого микробиологического производства является производственное культивирование соответствующего микроорганизма, проводимое либо с целью увеличения микробной массы - биомассы, либо для получения продуктов метаболизма растущей популяции микроорганизмов.
Под биомассой понимают общую массу особей одного вида, группы видов или сообщества микроорганизмов в целом. Ее выражают в массе сырого или сухого вещества (г/м2, г/м3). Отсюда ясно, что задача инженера-технолога - создание условий, обеспечивающих максимальную утилизацию компонентов питательной среды и накопление целевого продукта с заданными свойствами. Естественно, теоретической основой для этого являются закономерности, определяющие рост популяции микроорганизмов в зависимости от условий его осуществления. Знание количественных закономерностей роста популяций микроорганизмов в реальных условиях его осуществления в емкостной аппаратуре, выраженных в виде соответствующей математической модели, во многом определяет переход от эмпирического поиска к строгому решению задачи оптимизации технологических режимов получения продуктов микробиологического синтеза.
Периодическое культивирование
В процессе культивирования микроорганизмов периодическим способом, как указывалось ранее, можно выделить несколько периодов роста (рис. 2.19).

В первый период, после внесения в среду посевного материала (лаг-фаза), происходит процесс приспособления посевной культуры к новой среде. Численность популяции в эта время не увеличивается (а в некоторых случаях даже снижается). Состояние популяции в лаг-фазе формально можно описать так:
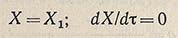
(для т, лежащего между 0 и т1).
Предполагается, что в период лаг-фазы микробные клетки не потребляют субстрата, но метаболическая активность клеток проявляется в повышении содержания белка и РНК (при постоянстве содержания ДНК), а также в увеличении объема клеток, который в общем виде может быть выражен с помощью уравнения
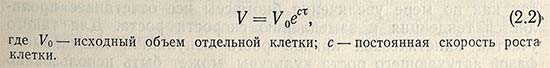
По достижении определенных соотношений между величинами поверхности клетки и ее объема происходит деление клетки, вследствие чего численность популяции начинает увеличиваться с возрастающей скоростью, которая для данной фазы роста культуры, называемой переходной, описывается соотношением
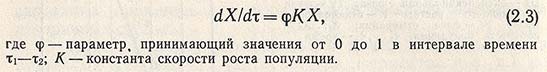
Интегральная зависимость, описывающая участок кинетической кривой роста между т1 и т2, имеет вид
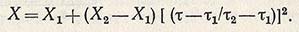
Увеличение скорости роста популяций в переходной фазе идет до предела, определяемого формально достижением параметром ф величины, равной единице, после чего скорость роста начинает выражаться зависимостью
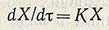
(для т между т2 и т3), откуда интегральная форма представляет экспоненциальную функцию
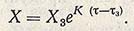
Эта фаза роста носит название экспоненциальной, или фазы логарифмического роста. Для оценки скорости роста биомассы часто пользуются величиной удельной скорости роста u.
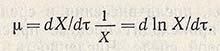
В качестве характеристики растущей культуры, находящейся в этой фазе, используют термины «время удвоения» и «время генерации» q, рассчитываемое по уравнению
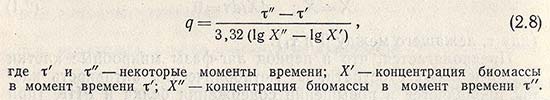
Однако такой характер роста популяции, который в первом приближении может быть описан экспоненциальной зависимостью, наблюдается в течение ограниченного периода времени, так как по мере увеличения биомассы все отчетливее проявляется тенденция к замедлению скорости роста. Для такого участка кинетической кривой роста популяции, называемого фазой затухающего роста культуры, может быть использовано дифференциальное уравнение для скорости роста
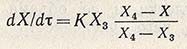
и его интегральная форма для описания изменения концентрации биомассы во времени
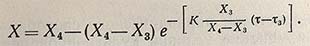
(для интервала времени между т3 и т4).
Снижение скорости роста по мере приближения X к значению Х4 происходит вплоть до достижения нулевого значения, которое характеризует вступление популяции в стационарную фазу: X = X4
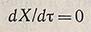
(для т, лежащих между т4 и т5).
По завершении фазы стационарного роста начинается фаза отмирания, или фаза дегенерации, культуры, характеризующаяся уменьшением численности популяции.
Приведенная выше система уравнений может быть использована только для описания конкретной кинетической кривой роста, полученной в результате эксперимента, но не в состоянии служить основой для прогнозирования процесса, так как в приведенных зависимостях в качестве параметров (Х1, ..., Х4; т1, ..., т5) вводятся конечные значения концентрации биомассы и времени. В настоящее время еще нет общепринятой математической модели роста популяции, которая достаточно точно описывала бы кинетику накопления биомассы в условиях периодического культивирования и содержала бы минимальное число эмпирических коэффициентов. В наибольшей степени этим требованиям отвечает модель Н. И. Кобозева, использование которой при изучении кинетики роста популяции дает обнадеживающие результаты. Интегральная форма предложенного им уравнения, описывающего кинетическую кривую роста популяции, имеет вид

Это уравнение является наиболее общим выражением для роста популяции, и в зависимости от частных условий (обратимое или необратимое размножение, рост популяции с исчерпыванием субстрата или при поддержании его количества на постоянном уровне) уравнение приобретает соответствующую форму и дает различное выражение для величины концентрации биомассы.
Основной недостаток периодического способа - цикличность и постоянное изменение условий культивирования, что затрудняет контроль и регулирование параметров процесса.
Большие возможности для повышения эффективности производства заложены в непрерывном способе выращивания.
Непрерывное культивирование
Сущность метода заключается в поддержании постоянных условий среды, а тем самым и микроорганизма-продуцента в определенном физиологическом состоянии. При непрерывном методе в образовании конечного продукта в течение всего ферментационного процесса участвует практически вся популяция микроорганизмов, чему способствуют оптимальные условия культивирования.
При непрерывном культивировании возникает открытая динамическая система, в которой микроорганизмы непрерывно размножаются со скоростью, зависящей от притока питательных веществ и других условий питания. Часть объема культуральной жидкости непрерывно вытекает с той же скоростью, с какой подается среда в аппарат, и количество микроорганизмов, поддерживающее непрерывный процесс, остается в ферментаторе постоянным. В стерильных условиях непрерывный метод обеспечивает сохранение культуры в физиологически активном состоянии длительное время.
Скорость увеличения биомассы в протоке выражается уравнением
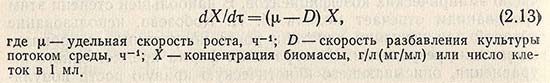
Значение D=F/V известно как скорость разбавления. Оно характеризует скорость потока на единицу объема. F - скорость протока среды, мл/ч (м3/ч); V - объем ферментатора, мл (м3).
Если в стационарном состоянии dX/dт = 0, то u = D. Это означает, что концентрация клеток неизменна. Чаще всего это бывает при D = 0,01 ± 0,25.
В условиях непрерывного культивирования, когда культура находится в состоянии динамического равновесия (при u = D), различают турбидостатный и хемостатный процесс.
При турбидостатном культивировании скорость протока среды регулируют так, что концентрация клеток остается постоянной. При хемостатном культивировании постоянная концентрация клеток в среде поддерживается при помощи постоянной концентрации химических соединений, в частности лимитирующего субстрата (например, источников углерода, азота, витаминов и др.).
Зависимость удельной скорости роста культуры от концентрации субстрата определяется уравнением Моно

В полноценной питательной среде Ks по отношению к S - величина незначительная и ею можно пренебречь, тогда из уравнения u = uмакс S/S = uмакс видно, что в полноценной питательной среде удельная скорость роста культуры не зависит от концентрации лимитирующего фактора. Для бактерий, растущих в питательной среде с углеводами, величина Ks составляет несколько десятых миллиграмма на 1 л среды, а для микроорганизмов, растущих на средах с аминокислотами, несколько микрограммов на 1 л, а 5 составляет несколько граммов на 1 л. Низкие значения Ks получены для дрожжей, выращиваемых на глюкозе. Следует отметить, что значение S/(Ks+S) близко к единице до тех пор, пока концентрация субстрата не слишком мала. Ввиду того что необходимая концентрация источника углерода для большинства ферментационных процессов выражается в г/л, то Ks в 10-100 мг/л сахара может вызвать снижение удельной скорости роста только на несколько процентов по сравнению с uмакс. В связи с этим величина удельной скорости роста в этих условиях не должна снижаться ниже 90% максимальной скорости роста.
Концентрация субстрата не является единственным фактором, лимитирующим скорость роста микроорганизмов. Н. Д. Иерусалимский пришел к заключению, что удельная скорость роста зависит от плотности популяции и что при высокой концентрации клеток задерживать рост могут продукты обмена веществ. Удельную скорость роста можно рассчитать по уравнению

Для поддержания культуры в состоянии максимальной скорости размножения необходимы постоянный приток свежего субстрата и вывод продуктов обмена веществ.
При определении прироста микроорганизмов в период культивирования предполагается, что содержимое ферментатора хорошо аэрируется и перемешивается, что популяция микроорганизмов однородна и свойства ее практически постоянны при высокой концентрации субстрата, лимитирующего рост, т. е. при S > Ks. Другие вещества, влияющие на рост, также находятся в постоянном избытке. Тогда удельную скорость роста u следует считать близкой к uмакс.
При периодическом культивировании микроорганизмов зависимость изменения концентрации микроорганизмов во времени описывается дифференциальным уравнением

Экономический коэффициент выражает количественные потребности микроорганизмов в питательных веществах. Если система находится в равновесии, то u = D. Равновесие можно нарушить, изменяя скорость протока u > D или u < Д или изменяя концентрацию субстрата среды S. Так как выход Y определяется из соотношения между образовавшейся биомассой и потреблением субстрата, то уравнение принимает вид
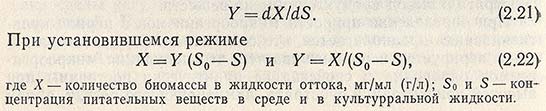
Следует отметить, что продуктивность процесса определяется произведением скорости разбавления на концентрацию микроорганизмов DX, т. е. количеством биомассы, получаемой с единицы объема ферментатора в единицу времени. Максимальная продуктивность процесса всегда связана с повышенной концентрацией субстрата в вытекающей среде.
Продуктивность непрерывного процесса выращивания может быть выражена уравнением
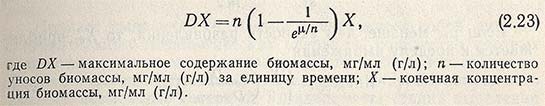
Сравнивая продуктивность периодического и непрерывного процессов культивирования в производстве кормовых дрожжей, предположим, что периодический процесс закончится за 10 ч, поэтому n = 1/10. Для выбранной величины u = 0,3 получаем D = 1/10 (1-1/3) = 0,095. При непрерывном производстве D = 0,3. Из этого следует, что при одинаковой концентрации микроорганизмов продуктивность процесса периодического культивирования втрое ниже, чем непрерывного. Когда цель культивирования - получение продуктов метаболизма Р, для описания процесса и определения биосинтетической активности К микроорганизма используют уравнение:
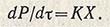
Это свидетельствует о том, что скорость биосинтеза метаболита за время dт прямо пропорциональна количеству биомассы и удельной активности К микроорганизма. К можно определить по уравнению

При непрерывном культивировании могут образовываться мутации. Если количество мутаций в течение одной генерации выразить через Q, число генераций в час D/ln2, удельную скорость размножения мутантов через uх и их концентрацию в ферментаторе через Хm, то уравнение примет вид
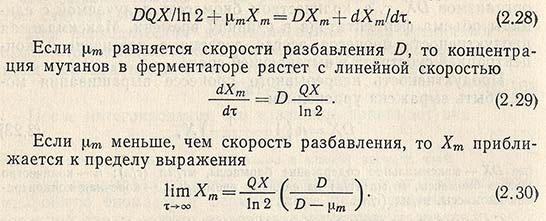
По этому уравнению можно вычислить концентрацию мутантов в ферментаторе.
В основе биологического синтеза лежат ферментативные процессы. Зависимость скорости ферментативной реакции от субстрата выражается согласно уравнению Михаэлиса-Ментен
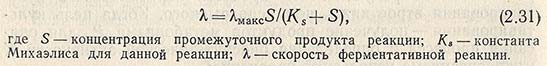
При непрерывном процессе u = D. Для образования продукта необходимо установить лимитирующий фактор и факторы, ограничивающие рост культуры (температура, pH, репрессоры, ингибиторы и т. д.).
При гомогенно-непрерывном процессе, где u = D
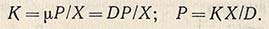
Из этого следует, что количество продукта зависит от концентрации биомассы, активности культуры и скорости разбавления D.
Материальный баланс гомогенно-непрерывного процесса по субстрату выражается уравнением
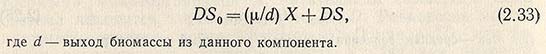
Можно допустить, что выход биомассы не зависит от концентрации компонента. Тогда по уравнению S = S0 – X/a можно определить концентрацию субстрата в вытекающей культуральной жидкости. С уменьшением величины скорости разбавления D увеличивается разница между концентрацией субстрата в притоке и оттоке, так как возрастает концентрация биомассы в среде.