Процессы питания и дыхания у микроорганизмов
Микроорганизмам, как и любым другим живым существам, свойственна одна и та же общая закономерность - обмен веществ с окружающей средой. Сущность процесса обмена у микроорганизмов состоит в том, что микробная клетка все время получает из субстрата питательные вещества. Эти вещества внутри клетки претерпевают различные превращения. Часть их тратится на построение новой живой цитоплазмы и других составных частей тела; часть же выделяется обратно во внешнюю среду как ненужные продукты обмена.
Совокупность тех процессов жизнедеятельности организма, которые связаны с обменом веществ и приводят к накоплению живой массы клеток, принято называть физиологическим процессом питания. С энергетической точки зрения питание любого организма нужно рассматривать как процесс накопления энергии, аккумулирование которой происходит в свойственных данному организму сложных пластических материалах - белках, жирах, углеводах.
Усвоение (ассимиляция) веществ пищи и превращение их путем сложных химических реакций в вещества, характерные для живой цитоплазмы данной клетки, выполнение всех других жизненных функций (движение, размножение и пр.) не может осуществляться клетками без затраты определенного количества энергии. Эту энергию живые существа получают в процессе дыхания. При дыхании каждое мгновение и непрерывно на протяжении всей жизни организма некоторая часть его клеточного вещества или сложных веществ, непосредственно им воспринятых, подвергается распаду (диссимиляции) на менее сложные соединения. Энергия, высвобождающаяся при этом, и используется клетками для выполнения их жизненных отправлений.
Процессы питания и дыхания неотделимы один от другого, взаимозависимы и осуществляются в микробных клетках одновременно. В процессе питания и дыхания происходят те сложнейшие качественные и количественные химические превращения органических веществ, которые и обусловливают обмен веществ живой клетки, а следовательно, и ее способность к жизнедеятельности.
Химический состав микробной клетки
Для различных видов микробов количественные соотношения и химическая структура питательных веществ должны быть, строго определенны - они зависят от химического состава клеточного вещества микроба. Химический состав микробных клеток в основном тот же, что и у клеток высших растений и животных. Главную массу живой цитоплазмы составляет вода: 80-85% ее массы. Остальные 15-20% приходятся на долю сухих веществ. Содержание воды в цитоплазме различных микробов неодинаково. Оно зависит от особенностей данного вида микроба, его возраста, питания и состава среды.
В воде растворены все наиболее важные органические и минеральные вещества, входящие в состав тела микробной клетки; в водной среде протекают основные биохимические процессы - такие, как гидролитическое расщепление белков и углеводов, в которых вода принимает участие как компонент. В цитоплазме у кишечной палочки содержится, например, 73% воды, у протея - 80%, в клетках плесневых грибов содержание воды доходит до 85%, а у дрожжей колеблется в пределах: 68-83%.
В состав сухих веществ микробной клетки входят: органогены - углерод, водород, кислород и азот, составляющие 90-97% сухого веса, и минеральные (зольные) элементы, на долю которых приходятся остальные 3-10% сухих веществ. В золе микробных клеток были обнаружены в очень незначительных количествах различные микроэлементы - бор, молибден, медь, цинк, бром, йод и некоторые другие. Органогены идут на построение сложных органических соединений: белков, жиров и углеводов.
Белки являются важнейшей составной частью органического вещества микробов. В состав тела микробных клеток входят простые и сложные белки. Наиболее важными для: жизнедеятельности микробов являются сложные белки, в частности: нуклеопротеиды, состоящие из нуклеиновых кислот, соединенных с белковой частью молекулы. Нуклеопротеиды содержатся главным образом в ядерном веществе клеток. Встречаются в составе цитоплазмы и глюкопротеиды; обнаружены они также в составе оболочки и бактериальных слизей.
Жиры и жироподобные вещества (липиды) находятся главным образом в оболочках клеток и в поверхностном слое цитоплазмы. Содержание жировых веществ в зависимости от вида микроба колеблется от ничтожно малых количеств до 30-60% сухих веществ; в среднем оно составляет 1-4%, но некоторые дрожжеподобные и кислотоустойчивые микроорганизмы накапливают жира значительно больше. Туберкулезная палочка, например, может содержать 20-30% жира, дрожжеподобный организм Endomyces vernalis («жировые дрожжи») накапливает до 30-47%.
Из углеводов в микробных клетках чаще всего встречаются полисахариды и продукты их гидролиза. В цитоплазме многих микробов в виде запасных питательных веществ, как уже указывалось, накапливаются зерна гликогена, декстрина, у некоторых обнаружены глюкуроновая кислота и глюкоза. Углеводы входят в состав оболочек и капсул микробных клеток, участвуют в синтезе белков и жиров и являются основным энергетическим материалом, расходуемым в процессах дыхания микробных клеток.
Минеральные вещества играют важную роль в регулировании осмотического давления внутри клетки, обусловливают благоприятное течение биохимических реакций. Наиболее важными зольными элементами являются фосфор, калий, кальций, железо и т.д. Микроэлементы: цинк, медь, бор, молибден и др. - играют важнейшую роль в процессах жизнедеятельности клетки.
Живая микробная клетка имеет определенную и обычно постоянную концентрацию ионов водорода в цитоплазме независимо от того, находится ли она в кислой или щелочной среде. Реакция цитоплазмы у микроорганизмов слабощелочная, а электрический заряд у большинства из них отрицательный. Относительная плотность микробных клеток, как правило, больше единицы (1,04-1,03), причем молодые культуры имеют несколько большую плотность, чем старые, но у вегетативных клеток микробов плотность меньше, чем у спор.
Изучение химического состава микроорганизмов имеет большой практический интерес. В некоторых отраслях нашего народного хозяйства применяются отдельные виды микроорганизмов, накапливающих в качестве запасных питательных веществ белки и жиры (например, кормовые и жировые дрожжи). Дальнейшие работы в этой области позволят выявить новые возможности использования полезной деятельности микробов и будут способствовать разработке новейших мер борьбы с вредными микроорганизмами.
Поступление питательных веществ в микробную клетку
Большинство микробов - одноклеточные растительные организмы. В отличие от высших растений многие из них не содержат хлорофилла - зеленого пигмента, в присутствии которого под влиянием солнечной энергии в листьях происходит синтез органических веществ. Нет у них и специальных органов пищеварения, имеющихся у высших животных. Одноклеточная структура многих микроорганизмов обусловливает характерные особенности процесса их питания.
Для осуществления процессов питания микроорганизмам необходима вода. Питательные вещества поступают в микробную клетку в виде растворов, в растворенном же виде удаляются из клетки отработанные, иногда исключительно сложные продукты обмена. Вода является главнейшей составной частью цитоплазмы: она входит в состав молекул белков, жиров, углеводов и продуктов их распада. Вода принимает непосредственное участие в огромном количестве химических, физико-химических и биохимических реакций, протекающих в живой клетке. Наличие значительного количества воды в цитоплазме необходимо для нормального протекания в ней жизненных процессов, так как обмен веществ с окружающей средой может происходить только при посредстве воды.
Процесс обмена между микробной клеткой и внешней средой происходит очень быстро и интенсивно, потому что поглощение питательных веществ и выделение в субстрат продуктов жизнедеятельности совершаются у микробов всей поверхностью их клеточной оболочки.
В основе механизма проникновения питательных веществ сквозь клеточную оболочку лежат сложные физико-химические явления - диффузия, осмос и обменная адсорбция. Считается, что протекают они параллельно. Однако в зависимости от температуры, концентрации ионов водорода в растворе (pH среды), концентрации и химической природы питательных веществ и некоторых других факторов одно из этих явлений может преобладать, хотя два других при этом не исключаются. Таким образом, питание микроба осуществляется комплексно всеми возможными путями.
Слово «диффузия» (diffusio) по-латински означает распространение. Диффузией называют взаимное проникновение частиц одного вещества между частицами другого, обусловленное их хаотическим движением. Основываясь на явлении диффузии, механизм проникновения питательных веществ в микробную клетку упрощенно можно было бы представить следующим образом. Вследствие того, что частицы вещества во всем объеме раствора стремятся выровнять свою концентрацию, возникает диффузный поток питательных веществ внутрь микробной клетки. Интенсивность этого потока поддерживается тем, что внутри клетки нужные для нее вещества быстро «усваиваются» - химически преобразуются (трансформируются) в вещества клетки микроба. Благодаря этому концентрация нужных для клетки веществ в поверхностном слое цитоплазмы все время оказывается низкой, обусловливающей непрерывную диффузию их из окружающей среды к поверхности тела микроба. Бесполезные или вредные для микроорганизма вещества химически не преобразуются клеткой, и потому концентрация их как внутри клетки, так и снаружи в окружающей среде быстро выравнивается: устанавливается подвижное диффузионное равновесие и поступление вредных веществ внутрь клетки прекращается.
Однако на самом деле поступление различных веществ сквозь клеточную оболочку значительно сложнее и во многих случаях подчиняется законам осмоса. Слово «осмос» (osmos) по-гречески означает «толчок». Осмос - явление, наблюдаемое, когда в соприкосновение приходят два раствора разной концентрации, отделенные один от другого полупроницаемой перегородкой (мембраной). Мембрана пропускает частицы чистого растворителя, но препятствует прохождению частиц растворенного вещества. Растворитель проникает через полупроницаемую мембрану в обе стороны; однако если с одной стороны мембраны находится более концентрированный раствор, то в эту сторону поступление молекул растворителя окажется гораздо большим, чем в обратном направлении. Идущая с количественным перевесом в одну сторону диффузия растворителя продолжается до тех пор, пока концентрации растворов не выровняются и не установится подвижное равновесие. То количество растворителя, которое переходит в раствор с более высокой концентрацией, оказывает на полупроницаемую мембрану определенное давление. Величина этого давления (его принято называть осмотическим) будет тем больше, чем выше разность концентраций между растворами, находящимися по обе стороны мембраны.
Полупроницаемые мембраны живых организмов (а таковыми являются стенки кишок, мочевого пузыря, оболочки растительных клеток, в том числе и микробных и пр.) обладают очень мелкими порами и не являются идеальными мембранами, пропускающими только растворитель - воду. Вместе с водой внутрь живой клетки проникают и те «частицы», диаметр которых меньше диаметра пор мембраны. Так, внутрь микробной клетки через клеточную оболочку очень быстро проникают углеводы (например, глюкоза) и другие соединения, не распадающиеся на ионы с малой молекулярной массой. Проникновение электрически заряженных частиц и ионов внутрь микробной клетки оказывается более сложным.
Выше указывалось, что бактериальные клетки в питательных субстратах чаще всего несут отрицательный заряд. Коллоидные частицы питательных веществ и ионы различных минеральных солей, имеющие заряд, одинаковый с оболочкой микробной клетки, вследствие электростатического отталкивания не могут проникнуть внутрь клетки микроба. Частицы же и ионы с противоположным знаком свободно проходят сквозь «поры» оболочки, если этому не препятствуют их размеры. Полисахариды и белки, коллоиднорастворимые в воде, имеющие очень большие размеры молекул, не могут проникнуть в клетку до тех пор, пока не подвергнутся гидролитическому расщеплению. Это расщепление осуществляется с помощью особых веществ - ферментов, выделяемых микробами наружу в питательную среду. Возникающие при гидролизе водорастворимые продукты уже значительно легче проникают (диффундируют) в микробную клетку.
Изучение свойств клеточной оболочки микроорганизмов показало, что это в высшей степени сложная коллоидно-дисперсная система. Наиболее существенной особенностью клеточной оболочки микробов - так называемой цитоплазматической мембраны - является ее постоянная изменчивость, или лабильность (от латинского labilis - нестойкий, неустойчивый). Цитоплазматическая мембрана имеет «мозаичную структуру». Отдельные ее участки имеют различную химическую природу: один участок представлен жироподобными веществами (липидами), другой - протеинами (белковоподобными веществами), третий состоит из веществ, имеющих углеводный характер, и т.д. Поэтому различные вещества могут проникать сквозь клеточную оболочку как благодаря растворению в липидах, так и путем «фильтрации» через водно-протеиновые поры между отдельными частицами оболочки. Чем крупнее молекула питательного вещества, тем труднее она проникает через поры цитоплазматической мембраны. Однако прямой зависимости здесь нет, так как цитоплазматическая мембрана обладает еще и свойством адсорбции.
Адсорбция - это поверхностное поглощение. Из окружающей среды клеточная оболочка способна «поглощать» (притягивать к своей поверхности) молекулы и ионы питательных веществ. Гораздо лучше адсорбируются клеточной оболочкой те вещества, которые уменьшают поверхностное натяжение (или положительно адсорбируются). Положительно адсорбирующиеся вещества, даже с очень крупной молекулой, могут свободно проходить через поры клеточной оболочки. Кроме того, в процессе дыхания в живой клетке постоянно образуется углекислый газ. При растворении в воде он образует угольную кислоту, диссоциирующую на ионы:
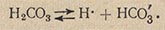
Ионы Н- и НСО'3, достигая клеточной оболочки, могут стать ее временными компонентами и легко обмениваются оболочкой на другие катионы и анионы питательной среды. Такое явление называется обменной (или полярной) адсорбцией.
Как видим, механизм проникновения питательных веществ внутрь микробной клетки оказывается очень сложным. Если учесть, что заряд клеточной оболочки зависит от pH питательного субстрата и при определенном значении его может измениться, что растворимость веществ и скорость диффузии, осмоса, адсорбции зависят от температуры, что концентрации веществ также влияют на протекание этих физико-химических процессов, то вполне понятным будет требование создания определенных условий при культивировании микробов. Изменение привычных условий жизни обусловливает у микроорганизма изменение проницаемости клеточной оболочки, что резко сказывается на протекающем обмене веществ и может либо вызвать гибель микроба, либо, наоборот, способствовать активному его развитию.
Важнейшим условием для нормального протекания обмена веществ, а следовательно, для нормального роста и размножения микробов является определенное напряжение клеточной оболочки, получившее название тургора. В состоянии тургора микробные клетки находятся в питательных средах, - концентрация веществ в которых несколько ниже, чем концентрация растворенных веществ внутри микробной клетки. В этом случае поступающая из питательной среды вода вызывает некоторое повышение осмотического давления клеточного сока и небольшое набухание цитоплазмы, в результате чего цитоплазма плотно прижимается к оболочке. Только при наличии тургора в клетках быстро и нормально протекают все процессы обмена веществ.
Если же микробная клетка попадет в среду со слишком высокой концентрацией веществ, то цитоплазма клетки начинает обезвоживаться. Вода из цитоплазмы уходит в субстрат, цитоплазма сжимается в комочек и постепенно отрывается от оболочки. Клетка приходит в состояние вялости. Наступает явление плазмолиза. При длительном плазмолизе микроорганизмы погибают. Явление плазмолиза лежит в основе методов консервирования пищевых продуктов концентрированными растворами соли или сахара (засол мяса, рыбы, приготовление варенья, засахаривание фруктов), а также методов сушки.
И, наконец, если микробная клетка попадает в среду с очень низкой концентрацией питательных веществ или в дистиллированную воду, то вследствие слишком высокой разницы в концентрации веществ в клетке и снаружи вода с силой и в большом количестве устремляется в клетку, вызывая чрезмерное набухание цитоплазмы. Цитоплазма растягивает клеточную оболочку до крайних пределов и даже может вызвать ее разрыв. Это явление носит название плазмоптиса. При плазмоптисе микроорганизмы также погибают.
Ввиду плазмоптиса микробных клеток микробиологи в своей практической работе не должны пользоваться дистиллированной водой при разведении культур микробов или при любых других: работах с живыми микробами (например, при получении смывов с поверхности исследуемых объектов и пр.).
Классификация микроорганизмов по способам питания
По способам питания, по условиям существования и по тем изменениям, которые они производят в среде своего обитания, микроорганизмы чрезвычайно сильно отличаются друг от друга. Это различие между микроорганизмами определяется прежде всего их способностью усваивать углеродистые и азотистые соединения.
Выше указывалось, что в процессе обмена веществ через цитоплазменную оболочку внутрь микробной клетки в преобладающем количестве поступают те соединения, из которых микроорганизм синтезирует собственное клеточное вещество и которые он использует в качестве дыхательного материала.
Микроорганизмам, как и другим живым существам, необходимы следующие элементы питания: углерод, азот, кислород, водород, сера, фосфор и другие зольные элементы, а также дополнительные факторы роста типа витаминов, микроэлементов и пр. Водород и кислород в случае необходимости микроорганизмы могут получить и из воды, а кислород некоторые микробы получают, кроме того, еще и из воздуха. Требования к источникам минеральных элементов у большинства микроорганизмов более или менее одинаковы. Многие микроорганизмы способны усваивать серу из сернокислых солей; лучшим источником фосфора являются соли ортофосфорной кислоты; калий, магний, кальций, железо микроорганизмы усваивают из их водорастворимых солей.
Микроэлементы, такие, как медь, бор, молибден, цинк, кобальт, марганец, ванадий и другие, сильно активирующие жизнедеятельность микробов и участвующие в синтезе ферментных белков, микроорганизмы могут усваивать из минеральных солей этих элементов или минеральных солей других питательных элементов, содержащих тот или другой микроэлемент в качестве примеси. Важно только, чтобы все эти соли находились в воде в нужном соотношении и не создавали неблагоприятной для микроорганизма реакции среды. Лишь некоторые микробы лучше усваивают серу из белковых веществ или из серусодержащих аминокислот (цистина, цистеина), а фосфор - из белков нуклеопротеидов.
Таким образом, особой разницы в питании микробов в смысле формы, в какой они усваивают зольные элементы, не наблюдается. Но к формам углеродистых и азотистых соединений, необходимым для питания, микроорганизмы предъявляют специфичные требования. По способности усваивать углеродистые соединения микроорганизмы подразделяются на следующие физиологические группы.
1. Автотрофные микроорганизмы (от греческого autos - сам и trophe - пища; иначе - «самопитающийся»). Они способны усваивать углерод непосредственно из углекислоты и не нуждаются в готовых органических соединениях. Составные части тела они могут синтезировать из СO2, Н2O и простых азотистых, соединений (аммиака, его солей, солей азотной кислоты). Углекислота для этих микроорганизмов служит единственным источником углерода. Более сложных соединений углерода автотрофные микроорганизмы усваивать не в состоянии.
К автотрофным микроорганизмам относятся нитрифицирующие бактерии, серобактерии, а также азотфиксирующие бактерии, способные усваивать свободный азот воздуха, водородные бактерии и железобактерии.
Синтез сложных, энергетически ценных органических веществ собственного тела из не имеющей энергетической ценности угольной кислоты автотрофные микроорганизмы осуществляют либо за счет энергии сопряженно протекающих химических реакций, либо за счет энергии солнечного света.
Способ получения энергии путем окисления H2S, NaNO2, NH4OH и других веществ, т.е. в результате химических реакций, был назван хемосинтезом, а микроорганизмы, которые обладают этой способностью, получили название хемосинтезирующих. Хемосинтез - совершенно исключительный физиологический процесс, представляющий огромный общебиологический интерес. Впервые хемосинтез был обнаружен выдающимся русским микробиологом С. Н. Виноградским в 1887 г. у нитрифицирующих бактерий. Нитрифицирующие бактерии неоднородны. Одна их группа - нитрозные бактерии - превращает аммиак в азотистую кислоту:

Вторая группа - нитратные бактерии - окисляют азотистую кислоту в азотную:
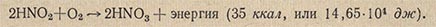
Обе группы нитрифицирующих бактерий часть химической энергии указанных реакций используют для синтеза органических веществ собственной природы:
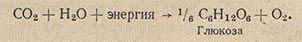
Процесс нитрификации - превращение аммиака в нитраты (селитры) - распространенный почвенный процесс, способствующий накоплению в почве селитр - важнейших питательных веществ для растений. Поэтому процесс нитрификации имеет большое значение в сельском хозяйстве. У бесцветных серобактерий источником энергии для синтеза клеточного вещества является реакция окисления сероводорода до серной кислоты, у тионовых бактерий - окисление до серной кислоты тиосульфитных (Na2S2O3) и тетратионовых (Na2S4O6) соединений и т.д.
Фотосинтезирующие автотрофные микроорганизмы - это микробы, которые необходимую для синтеза энергию получают в виде энергии солнечных лучей. Процесс синтеза сложных органических соединений за счет энергии солнца, как известно, называется фотосинтезом. Фотосинтезирующими являются пигментные бактерии, имеющие в своих клетках пигменты (красящие вещества) типа хлорофилла, например зеленые и пурпурные серобактерии. В клетках пурпурных серобактерий содержится красное (или пурпурное) вещество - бактериопурпурин, по химическому составу близкое к каротиноидам (группа ликопина). В теле зеленых (или сине-зеленых) серобактерий содержится зеленый пигмент - бактериохлорофилл (или бактериохлорин) и хлоробиум-хлорофилл, образующие при окислении продукты, подобные продуктам окисления хлорофилла. Имеются в теле сине-зеленых серобактерий и каротиноидные пигменты. Роль красящих пигментов в теле фотосинтезирующих микробов такая же, как и у хлорофилла зеленых листьев: эти пигменты участвуют в поглощении световой энергии, т.е. в том процессе, который составляет основу фотосинтеза. Порчи пищевых продуктов автотрофы не вызывают.
2. Гетеротрофные микроорганизмы (от греческого heteras - иной, trophe - пища; дословно - «питающийся иным»). Эти микроорганизмы для своего питания нуждаются в органическом углероде (углеводы, кето-, амино-, окси-, жирные кислоты), а также в различных азотистых соединениях (нитраты, аммиак), в минеральных веществах, микроэлементах и витаминах.
Многие гетеротрофы развиваются на самых разнообразных органических веществах независимо от их химической структуры, физико-химических свойств или реакционной способности. Но существуют и такие, которые нуждаются в строго определенном углеродсодержащем органическом веществе.
Гетеротрофов принято делить на сапрофитов и паразитов. Уже указывалось, что сапрофиты составляют большинство микроорганизмов, населяющих нашу планету. Это - возбудители гниения и самых разнообразных брожений. О грандиозной общебиологической роли сапрофитов, о их большом техническом значении подробно сказано во введении. Большинство известных в настоящее время бактерий, дрожжей, плесеней принадлежит именно к сапрофитам.
В группу паразитов входит сравнительно небольшое количество видов микробов, приспособившихся в ходе эволюции к паразитическому образу жизни. Они живут на поверхности или внутри другого организма-хозяина, питаются за его счет сложными органическими веществами живых тканей. Своим присутствием паразиты нарушают различные физико-химические и химические процессы, протекающие в организме тех существ, в тканях которых они живут. Эти нарушения представляют собой болезнь, от которой пораженный организм может погибнуть. Примерами болезнетворных (патогенных) бактерий являются брюшнотифозные палочки, туберкулезные, дифтерийные, возбудители столбняка, сибирской язвы и пр. Так как эти микроорганизмы в зависимости от условий ведут себя то как паразиты, то как сапрофиты, их называют факультативными паразитами. Однако есть микроорганизмы, которые полностью утратили способность к сапрофитному образу жизни и могут жить только как паразиты внутри других живых клеток. Такие микробы получили название облигатных (безусловных) паразитов. Безусловный паразитизм наблюдается, например, у вирусов и некоторых патогенных простейших. Они не могут расти на искусственных питательных средах, на которых культивируются сапрофиты. Развиваясь за счет живых организмов, вирусы являются возбудителями многих инфекционных заболеваний человека, животных и растений.
И в то же время известны случаи, когда, казалось бы, заведомый сапрофит также вызывает заболевания человека или животных. Примером может служить кишечная палочка (Escherichia coli) - нормальный обитатель толстого отдела кишечника. Являясь сапрофитом (комменсалом) кишечная палочка в случае ослабления организма человека может вызвать заболевание, колиэнтеритом (воспаление кишечника), перитонитом (воспаление брюшины), пиелитом (воспаление почек). Таким образом, нельзя провести резкой границы между сапрофитами и паразитами. Не существует такой границы и между автотрофами и гетеротрофами.
В 1921 г. русским исследователем А. Ф. Лебедевым были открыты микробы, считавшиеся гетеротрофами, но свободно включающие в свой обмен веществ минеральную угольную кислоту. Этот промежуточный метаболизм (от греческого metabole - перемена; дословно - обмен веществ) был обнаружен у пропионовокислых бактерий, усваивающих углекислый газ из бикарбонатов. В настоящее время установлено, что не лишены этой способности и многие гетеротрофы и даже патогенные микроорганизмы. Таким образом, абсолютных гетеротрофов не существует.
По характеру азотистого питания микроорганизмы также подразделяются на ряд групп: 1) фиксирующие азот атмосферы; 2) усваивающие минеральные формы азота (аминоавтотрофы); 3) развивающиеся в присутствии аминокислот или их смеси; 4) развивающиеся в белковых питательных средах. Две последние группы микробов получили название аминогетеротрофов.
По способности к синтезу сложных соединений микроорганизмы разделены на четыре группы.
1. Микроорганизмы, усваивающие углерод из углекислого газа и азот из неорганических соединений. Сюда отнесены хемоавтотрофные и фотоавтотрофные микроорганизмы.
2. Микроорганизмы, способные усваивать углерод из органических соединений углерода, а азот из его неорганических соединений (большинство сапрофитов).
3. Микроорганизмы, нуждающиеся для своего питания в органических соединениях углерода и усваивающие азот из аминокислот (кишечная палочка и другие комменсалы).
4. Микроорганизмы, нуждающиеся в сложных органических соединениях как углерода, так и азота, требующие одного витамина и более (патогенные бактерии).
В особых источниках энергии гетеротрофные микроорганизмы, как правило, не нуждаются. Многие из них, усваивая определенные органические вещества в качестве питательного материала, одновременно могут использовать их и в актах дыхания (или брожения) как энергетический материал. Но в некоторых случаях этого не наблюдается. Вещества, используемые микробом в качестве пластического материала, могут оказаться неподходящими для дыхания и наоборот. Подобное явление обнаружено у уксуснокислых бактерий. Окисляя при дыхании этиловый спирт, уксусные бактерии не могут использовать его в синтезе пластических материалов своего тела, а нуждаются в углеводах или других каких-либо источниках углерода.
Для нормального развития многим микроорганизмам, помимо углеродистых, азотистых соединений, зольных элементов, необходимы, как выше указывалось, минимальные количества особых веществ - стимуляторов или факторов роста, которыми в большинстве случаев являются витамины. Избыток витаминов задерживает рост микробов. Не нуждаются в ростовых веществах автотрофы, так как они способны их синтезировать.
Факторы роста играют роль катализаторов в биохимических процессах в клетке, а также являются структурными единицами при образовании некоторых ферментов. Для своего развития значительная часть микробов требует наличия биотина.
Способность микроорганизмов синтезировать те или иные витамины и их потребность в тех или иных ростовых веществах в настоящее время достаточно хорошо изучена. Для промышленного производства витаминов используют многих микробов: из грибка эремотециум вырабатывают витамин В2, из пропионовокислых бактерий и некоторых актиномицетов - витамин В12, из дрожжей - витамин В1, пантотеновую кислоту и биотин. Микроорганизмами пользуются при аналитическом определении в тех или иных питательных веществах отдельных витаминов. Показателем содержания витамина в среде является пышный или скудный рост или полное отсутствие роста соответствующего микроорганизма.
В заключение укажем, что химические соединения, слагающие тело микробов, предопределяют лишь элементарный состав питательных сред. Нередко для питания микробов нужны совсем иные химические вещества, чем те, которые обнаружены в составе их клеточного вещества, и далеко не каждый микроорганизм будет развиваться в субстрате, составленном из химических соединений, слагающих его клеточное вещество. Очень часто соединения, служащие для одних микробов прекрасным источником углерода, для других оказываются совершенно непригодными и даже ядовитыми. Более того, вещества, наиболее легко усваиваемые микробом в одних условиях существования, могут оказаться малопригодными при изменении этих условий. Поэтому в практической работе чрезвычайно важное значение имеет правильный выбор питательной среды, содержащей все необходимые компоненты для нормального развития того или иного вида микроорганизма.
Дыхание микроорганизмов
В процессе жизнедеятельности для осуществления роста, развития и размножения микробная клетка, как и всякая другая живая клетка, требует определенного количества тепловой энергии. Необходимую энергию микроорганизмы получают в процессе дыхания. Сущность дыхания заключается в окислении сложных органических веществ до более простых с выделением тепловой энергии, которая и используется микроорганизмами.
Как правило, дыхание протекает в присутствии кислорода воздуха, однако еще Пастером было установлено, что существуют микроорганизмы, которые могут свободно развиваться в бескислородной среде. Микроорганизмы, которые требуют в процессе дыхания широкого доступа воздуха, получили название аэробных. Типичными аэробами являются, например, плесневые грибы, уксуснокислые бактерии и пр.
Микроорганизмы, которые не нуждаются в кислороде, были названы анаэробными. Степень анаэробности микроорганизмов весьма различна. Существуют микроорганизмы, которые развиваются в отсутствие кислорода, но совершенно свободно живут и при его наличии. Такие микроорганизмы называют факультативными (безразличными, или условными) анаэробами. К факультативным анаэробам принадлежат, например, дрожжи, молочнокислые бактерии.
Микроорганизмы, для которых кислород не только не нужен, но вреден даже в незначительных количествах, называются облигатными (строгими, или безусловными) анаэробами. Облигатные анаэробы достаточно широко распространены в природе. Их можно обнаружить там, где нет свободного кислорода или приток воздуха затруднен (на дне илистых отложений в стоячих водоемах, в глубоких слоях почвы, загрязненной растительными или животными отбросами, в кучах мусора и пр.). Примерами облигатных анаэробов могут служить маслянокислые микроорганизмы, некоторые возбудители гниения и др. Микроорганизмы в большинстве случаев получают необходимую для жизнедеятельности энергию путем окисления углеводов и некоторых органических кислот.
В случае аэробного дыхания вещество полностью окисляется до своих конечных продуктов - углекислого газа и воды, при этом выделяется значительное количество тепловой энергии. Типичным примером реакции, протекающей при аэробном дыхании, служит уравнение

Способ получения энергии при анаэробном дыхании назван брожением. Энергетический эффект брожения всегда более низкий, чем: при аэробном дыхании, так как при брожении происходит неполное окисление органического вещества. Большая часть потенциальной энергии остается заключенной в конечных продуктах брожения. Примером одного из видов брожения может служить спиртовое брожение:
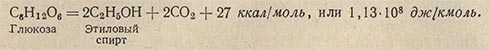
Количество энергии, выделяющейся при брожении, оказывается во много раз меньшим (почти в 28 раз), чем при дыхании. А так как потребность в энергии у различных микроорганизмов более или менее одинакова, то возбудителям брожения для получения всего необходимого количества энергии приходится перерабатывать гораздо больше вещества.
Брожение, таким образом, является менее совершенным методом добывания энергии, чем дыхание, и сопровождается огромной тратой дыхательного материала. Объяснить возникновение такого «неэкономного процесса» в результате эволюционного развития можно отсутствием необходимого кислорода. На дно стоячих водоемов кислород проникает с трудом, а именно на дне скапливается больше всего органических веществ. В таких условиях, очевидно, у организмов и выработалась способность добывать необходимую для жизнедеятельности энергию путем брожения.