Истинные грибы (плесени и дрожжи)
Обычно плесени и дрожжи рассматриваются как две отдельные группы грибов. Между ними обычно проводят морфологическое различие, определяя плесени как грибы, образующие ветвящиеся волокна (мицелий), а дрожжи - как одноклеточные грибы. Но истинного различия не существует, так как некоторые плесени принимают в определенных условиях дрожжеподобную форму, а многие дрожжи (обычно одноклеточные) могут образовать мицелий. Имеется определенное родство между дрожжами и высшими грибами. Дрожжи рассматриваются как примитивные формы. Тем не менее, для нашей цели удобнее рассматривать плесени и дрожжи отдельно.
Плесени
Обычные плесени состоят из трубчатых волокон микроскопического размера, называемых гифами, которые обильно ветвясь и переплетаясь, образуют паутинообразную структуру, называемую мицелием.
Во многих случаях мицелий может быть дифференцирован ка вегетативную часть, которая располагается на поверхности и внутри субстрата, на котором он прорастает, и плодоносящую спорообразующую часть, развивающуюся в воздухе. Гифы могут быть септированные или несептированные.
В первом случае каждая гифа разделена поперечными перегородками на клетки, каждая из которых содержит одно или несколько ядер. Несептированные гифы имеют вид удлиненных многоядерных трубок. Первоначальное деление гриба основано на наличии или отсутствии перегородки в гифе.
Удлинение гиф и рост мицелия происходят или на конце (верхушечный рост) или делением клеток в гифе (интеркалярный рост).
Размножение
Плесени нормально размножаются при помощи спор, которые обычно представляют собой маленькие тельца в несколько микрон в диаметре. Эти тельца образуются каждым организмом в большом количестве. Спорообразование у плесеней является истинным процессом размножения, так как каждая спора способна дать начало новым организмам - гифам, которые разрастаются и разветвляются, образуя свежий мицелий. При благоприятных условиях весь цикл от споры до новой споры может продолжаться весьма недолго (24-48 часов).
Идентификация плесеней базируется главным образом на особенностях их системы размножения, которая может быть по происхождению половой или бесполой. У многих видов имеются оба типа спорообразования, и в этих случаях половые споры рассматриваются как совершенная форма, наиболее важная для целей классификации; бесполые споры именуются несовершенными или добавочными, но их особенности и форма образования являются важным признаком в дифференциации видов даже для образующих половые споры.
Половой споруляции всегда предшествует слияние двух отдельных клеток гифа и их ядер; при бесполой споруляции споры образуются на концах плодоносящих гиф. У некоторых видов бесполые споры возникают одиночно или группами в результате почкования или сегментации плодоносящих гиф. Такие споры называются конидиями, а гифы, на которых они возникают, - конидиеносцами. Конидии могут прикрепляться к плодоносящей гифе короткими стебельками - стеригмами, у некоторых видов ветвящимися.
Несептированные плесени также могут образовывать внутри спорангия бесполые споры. Такие споры называются спорангиеспорами, а гифы, на которых они образуются - спорангиеносцами. Конец спорангиеносца, входящий внутрь спорангия, называется колумеллой, размеры и форма которой важны для целей классификации. Спорангии никогда не образуются септированными плесенями. У некоторых видов грибов гифы распадаются на короткие отрезки, каждый из которых рассматривается как спора, так как способен давать начало новому организму. Эти отрезки называются оидиями.
Бесполые споры, называемые хламидоспорами, образуются путем утолщения клеток гиф. Они возникают в любой части мицелия и не связаны с определенным положением. Эти споры обладают повышенной сопротивляемостью по отношению к воздействию неблагоприятных условий. У некоторых видов сплетения гиф образуют твердые компактные массы, именуемые склероциями, которые также способны противостоять неблагоприятным условиям.
Основные группы
Имеется много тысяч видов настоящих грибов, которые трудно классифицировать. Существует много различных систем, подробно описанных в руководствах. В пределах этой книги можно только сослаться на основные группы и указать важные роды и виды, вызывающие порчу пищевых продуктов. Истинные грибы обычно подразделяются на основании следующей схемы:
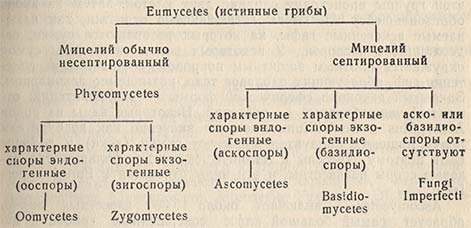
Phycomycetes
Этот класс делится на два подкласса - Oomycetes и Zygomycetes. Члены этой группы не представляют интереса с точки зрения порчи пищевых продуктов, но многие из них служат причиной заболевания растений и рыб.
У Zygomycetes половыми спорами являются зигоспоры. Они образуются в результате слияния двух гиф. В точке соприкосновения образуются выросты или прогаметангии, каждый из которых отделяется от родительской гифы образовавшейся перегородкой. Отделившиеся части сливаются, и стенка образовавшейся клетки становится толстой и бугристой. Эта группа делится на два порядка: Mucorales - бесполые споры в спорангии, Entomophthorales - бесполые споры на конидиеносцах, выбрасываемые при созревании.
Порядок Entomophthorales включает много видов, паразитирующих на насекомых, но их не обнаруживают в испорченных продуктах. Большинство видов Mucorales являются сапрофитами. Представители родов Mucor, Rhizopus и Thamnidium вызывают порчу пищевых продуктов.
Ascomycetes
Этот класс характеризуется образованием половых аскоспор внутри сумки или аскуса. Этот тип спорообразования часто называется эндогенным. Обычно в каждой сумке имеется восемь аскоспор, причем во многих случаях они могут быть образованы бесполым путем.
Способы образования сумок у разных видов сильно различаются между собой. У простейших форм, как например у дрожжей, первоначально происходит слияние двух смежных клеток и их ядер, после чего наступает деление с образованием Дочерних ядер, каждое из которых становится спорой. Таким образом, дрожжевая клетка образует сумку. У высших членов этой группы происходит слияние двух клеток, затем из вновь образовавшейся клетки вырастают новые отростки, так называемые аскогенные гифы, на которых развиваются сумки, содержащие аскоспоры. У некоторых видов скопление сумок окружается плотным защитным покровом, состоящим из сплетения гиф, образующих плодовое тело, называемое аскокарпом. Закрытый аскокарп сферической формы - это перитеций; открытый чашеобразный - апотеций. Некоторые виды из родов Aspergillus и Penicillium, имеющие значение как возбудители порчи пищевых продуктов, образуют сумки (аски) и должны быть поэтому отнесены к Ascomycetes, но для большинства половые споры неизвестны, и эти виды относятся к Fungi Imperfecti.
Ascomycetes - включает около 15 000 известных видов, образует самый большой класс совершенных грибов и делится на:
Plectomycetes - аскокарп отсутствует, а если и имеется, то он примитивный и неправильный с неравномерно расположенными асками.
Discomycetes - сумки расположены параллельными рядами в апотециях.
Pyrenomycetes - сумки расположены параллельными рядами в перитециях.
Plectomycetes - включает дрожжи и другие грибки, имеющие значение для промышленности. Среди Discomycetes имеются паразиты растений и виды сапрофитов; большие грибковые наросты на стволах деревьев, стенах и т. д. относятся к этой группе. Pyrenomycetes включает большое число вредителей растений, паразитов на насекомых и небольшое число обычных плесеней.
Из Ascomycetes, кроме дрожжей порчу консервированных продуктов вызывает Byssochlamys fulva. Аскоспоры этого вида необычайно термоустойчивы.
Basidiomycetes
Для этой группы характерны базидиоспоры, от которых она получила свое название. Этот класс включает шляпочные грибы (съедобные и поганки) и паразитические плесени. Он не имеет большого значения для пищевой микробиологии.
Fungi Imperfecti
Этот класс включает все виды, для которых неизвестно: половое или совершенное спорообразование: они размножаются бесполыми спорами. Большая часть обычных плесеней относится к Fungi Imperfecti и по количеству превосходит Ascomycetes. Известны три группы:
Sphaeropsidales - конидии образуются внутри бутылочковидных пикнидий.
Melanconiales - конидиеносцы образуются в блюдцевидном углублении на поверхности субстрата.
Hyphomycetales - свободные конидиеносцы поднимаются неравномерно из мицелия.
В группу Sphaeropsidales входит много видов паразитов на растениях. Группа Melaneoniales главным образом паразитирует на растениях и не имеет практического значения для консервированных продуктов.
Большинство сапрофитных плесеней, представляющих экономический интерес, относится к Hyphomycetales. Порчу пищевых продуктов вызывают роды: Aspergillus, Penicillium, Sporotrichum, Cladosporium, Alternaria, Fusarium, Oidium и Monilia.
Многие виды плесеней имеют большое промышленное значение. Примерами промышленного использования могут служить Aspergillus niger - для производства лимонной кислоты и Penicillium - для созревания некоторых видов сыров.
Большой интерес представляет Penicillium notatum - вид, образующий антибактериальное средство - пенициллин.
Дрожжи
Дрожжи не могут быть точно отделены от грибов, образующих типичный мицелий, однако Хенричи дал совершенно ясное определение термина дрожжи: это «настоящие грибы, у которых обычен и доминирует одноклеточный характер роста».
Дрожжевая клетка состоит из проницаемой клеточной стенки или мембраны, покрывающей цитоплазму, содержащую ядро и различные включения. У молодых клеток стенка тонка и цитоплазма сравнительно гомогенна, но при старении клетки происходит утолщение оболочки и в цитоплазме появляются включения и вакуоли. Ядро обычно незаметно, и его можно увидеть лишь в окрашенной клетке. Дрожжевые клетки, за исключением периодов активного размножения, отделены одна от другой и имеют, разнообразную форму, причем основными Формами являются круглая, овальная или эллипсоидная. Размер клеток - до 8 u в диаметре и до 10 u в длину.
Размножение
Размножение дрожжей происходит путем почкования, деления или спорообразования. Нормальным способом вегетативного размножения большинства важных для промышленности дрожжей является почкование. При этом процессе клеточная оболочка образует выпячивание, или почку, которая, суживаясь у своего основания и постепенно увеличиваясь примерно до размеров материнской клетки, отделяется от нее. Часто в еще не отделившихся почках образуются новые дочерние почки, и таким образом формируется группа связанных между собой клеток различной величины. При размножении делением клетка делится на две дочерние, как у бактерий.
В дополнение к этим вегетативным процессам размножение может происходить и путем образования спор. В одном семействе дрожжей встречается половое размножение, в котором клетка становится аскусом, содержащим до восьми аскоспор, и такие дрожжи подобны классу Ascomycetes грибов. Второе семейство имеет баллистоспоры, которые появляются на узких выростах, образуемых вегетативными клетками. Баллистоспоры выбрасываются таким же способом, как у группы Basidiomycetes. Можно было бы предположить родство между этими группами, но это вызывает сомнение. Дрожжи, не образующие аскоспор или баллистоспор, могут быть отнесены к несовершенным формам подобно несовершенным грибам.
Споры дрожжей более устойчивы к неблагоприятным условиям, чем вегетативные клетки, но, как и у плесеней, эта разница в устойчивости не так резко выражена, как у бактерий и их спор.
Классификация дрожжей
Дифференциация дрожжей основана главным образом на способах размножения, но одного этого недостаточно для разделения видов. Важными для диагностики являются биохимические реакции, культуральные признаки и другие особенности. Подробно технические приемы и среды, требуемые для идентификации дрожжей, даются в соответствующих руководствах. Имеется много систем классификации дрожжей. Самая поздняя различает три основные группы:
Endomycetaceae. Это семейство включает все виды, образующие аскоспоры.
Подсемейство I Eremaseoideae - только мицелий; вегетативное размножение путем деления клеток; аски образуются после конъюгации; спор до 8 в каждой аске; окислительная диссимиляция.
Род Eremascus.
Подсемейство II Endomycetoideae - мицелий и (или) артроспоры; вегетативное размножение путем деления клеток; образование аск обычно после конъюгации; споры круглые, овальные, цилиндрические; окисляют или ферментируют.
Род Endomyces, Schizosaccharomyces.
Подсемейство III Saccharomycetoideae - мицелий и почкующиеся клетки, псевдомицелий и (или) свободно почкующиеся клетки; вегетативное размножение посредством деления и почкования или только почкования; конъюгация может предшествовать образованию аск; споры различной формы, 1-4 в каждой аске; доминируют ферментирующие.
Подотряд A Endomycopseae.
Род Endomycopsis.
Подотряд В Saccharomyceteae.
Род Saccharomyces, Pichia, Hansenuia, Debaryomyces и Schwanniomyces.
Подотряд С Nadsonieae.
Род Saccharomycodes, Hanseniaspora и Nadsonia.
Подсемейство IV Nematosporoideae - мицелий и (или) почкование; вегетативное размножение путем деления клеток и (или) многостороннее почкование; конъюгация может предшествовать образованию аск; споры игольчатые или веретенообразные; сбраживают или окисляют.
Род Monosporella, Nematospora и Coccidiascus.
Подсемейство V Lipomycetoideae - только почкование; мешочковидные аски образуются вегетативными клетками в виде выростов; споры овальные, до 16 в каждой аске; сильные окислители.
Род Lipomyces.
Sporobolomycetaceae. Виды этого семейства образуют баллистоспоры, которые возвышаются на стеригмах и выбрасываются капельновыделительным механизмом.
Род Sporobolomyces и Bullera.
Cryptococcaceae - содержит все виды, не образующие ни аскоспор, ни баллистоспор.
Подсемейство I Cryptococcoideae - кроме почкующихся клеток образует псевдомицелий и настоящий мицелий, но не образует артоспор. Размножение главным образом почкованием, хотя может встречаться деление. Не образует ни красного, ни желтого пигментов.
Род Cryptococcus, Torulopsis, Pityrosporum, Brettanomyces, Candida, Kloeckera и Trigonopsis.
Подсемейство II Trichosporoideae - образует настоящий мицелий, псевдомицелий, артоспоры и почкующиеся клетки; размножение почкованием и делением; не образует ни красного, ни желтого пигментов; вообще окислители, но иногда ферментирующие.
Род Trichosporon.
Подсемейство III Rhodotbruloideae - образует почкующиеся клетки, псевдомицелий; размножение - многобоковое почкование; образует желтый до красного пигменты; окислители.
Род Rhodotorula.
Важные (для пищевой промышленности) роды
Saccharomyces. Этот род содержит много важных видов, из которых Saccharomyces cerevisiae (Hansen) наиболее известный. Образует спирт и углекислоту из сахара и широко применяется в пивоваренной и хлебопекарной промышленности. Размножается почкованием и бесполыми спорами. Другие важные виды этого рода: Saccharomyces cerevisiae вар. ellipsoideus (вообще известный как S. ellipsoideus). Этот вид используется для сбраживания вина. Вообще встречается на винограде и в почве виноградников. Клетки овальные или пирамидальные и обладают высокой устойчивостью к спирту, но, с другой стороны, подобны S. cerevisiae Hansen. Saccharomyces pasteurianus вызывает порчу пива и доставляет много забот пивоваренной промышленности.
Другие виды этого рода используются при изготовлении различных ликеров (яблочный сидр, грушевый сидр и др.) и сброженных молочных продуктов (кумыс, кефир и др.).
Saccharomyces rouxii и Saccharomyces mellix имеют важное значение в пищевой промышленности из-за их способности развиваться в продуктах с высокой концентрацией сахара, таких, как джем, мед, леденцовая пена и других. Многие штаммы ос мофильных дрожжей раньше относили к подроду Zygosaccharomyces. Леддер и Крегер-ван-Рий не признают этот подрод и относят его к роду Saccharomyces.
Pichiа и Hansenula. Эти роды представляют интерес как вредители пивоваренной промышленности. Они образуют эфиры (этилацетат, амилацетат и др.) из сахара, но не образуют спирта; в спиртовых жидкостях большинство видов используют спирт как источник углерода, и поэтому они вызывают серьезную порчу сбраживаемых ликеров. На жидкостях образуют сухие морщинистые пленки, часто обнаруживаемые на поверхности маринадов.
Debaryomyces. Виды этого рода обладают высокой устойчивостью к соли; обычно их обнаруживают в маринадах.
Torulopsis. Виды этого рода часто загрязняют пищевые продукты. Некоторые виды являются осмофилами и вызывают порчу меда.
Вretta nomyces. Этот род содержит дрожжи, имеющие значение в пивоваренной промышленности. Их обнаруживают в пиве после ферментации.
Candida. Различные виды, ранее отнесенные к роду Mycoderma, сейчас относят к единственной разновидности - Candidamycoderma, образующей сухие морщинистые пленки на жидкой среде. Спирт используют как источник углерода. Эти дрожжи часто вызывают порчу сбраживаемых напитков. Их также обнаруживают на маринадах. Candida utilis, ранее известный как Torula utilis, быстро растет в массе. Из-за высокого содержания белка и также витаминов его используют для питания животных и людей.
Rhodotorula. Все виды образуют красный, розовый, оранжевый или желтый пигмент каротиноидной природы. Сбраживаемая способность у них низкая. Загрязняют воздух и вызывают порчу пищевых продуктов. Согласно Педерсону и Келли, они являются причиной розовой окраски квашеной капусты.
Биологическая характеристика грибов
Грибы не содержат хролофилла и не могут питаться исключительно неорганическими веществами, поэтому все они гетеротрофы и для энергетических целей нуждаются в органическом источнике углерода, а некоторые - и в органическом, азоте. Для питания грибов имеют также значение элементы: калий, фосфор, сера, магний, и иногда железо. Эти элементы обычно используются грибами в виде неорганических соединений. Плесени отличаются способностью развиваться при минимальных Количествах пищевых веществ. Часто растут в неорганических солевых растворах, на поверхностях тканей, металла, стекла, где содержание питательных веществ очень низкое.
Грибы усваивают пищевые вещества подобно бактериям через клеточную стенку. Они обладают большим количеством разнообразных ферментов, что дает возможность различным видам разлагать углеводы, белки, жиры и органические кислоты. Основными продуктами, получаемыми в результате брожения, вызываемого грибами, являются нелетучие кислоты и спирт. Может продуцироваться и углекислота, но только не водород и не метан; в этом отношении грибы отличаются от бактерий, так как некоторые виды бактерий образуют водород и реже метан.
По своей потребности в кислороде большинство обычных плесеней строго аэробны, но некоторые виды способны расти при значительно пониженном давлении кислорода. Многие дрожжи являются факультативными анаэробами, но не существует действительно анаэробных грибов.
К температуре грибы относятся различно. Температурный оптимум роста для большинства грибов лежит между 25-30°. Многие виды не растут при температуре выше 30°, но для некоторых видов такая температура является оптимальной. Плесень Aspergillus furaigatus может существовать при температуре выше 50°, хотя спорообразование и оптимум роста наблюдаются при 40°. Кастелл наблюдал рост двух видов Mucor при 55°. Температурный оптимум для развития мицелия не должен обязательно совпадать с оптимумом для других процессов, например, полового спорообразования. Низший предел для роста также варьирует, и развитие некоторых видов, например Cladosporium herbarum, наблюдается при температуре около -6°.
Плесени также неодинаково стойки к нагреванию, причем споры несколько устойчивее, чем вегетативные клетки, но уступают в этом отношении бактериальным спорам. Некоторые виды разрушаются при длительном воздействии температуры 30°, а большинство видов разрушаются при 30-минутном нагревании при 65°. Исключением является плесень Byssochlamys fulva, вызывающая порчу консервированных фруктов. Споры этого вида выдерживают нагревание при 87-88° в течение 30 мин. Вильямс, Камерон и Вильямс сообщают, что склероции одного из видов Penicillium разрушались лишь при нагревании в течение 300 мин. при 85°.