Влияние нагревания на микроорганизмы
Знание основных принципов и различных факторов, влияющих на уничтожение микроорганизмов нагреванием, имеет существенное значение для полного понимания задач, возникающих в связи с тепловой стерилизацией консервов. В этом направлении проделана большая работа, в результате которой изучены различные методы тепловой стерилизации; все же остается еще много вопросов, подлежащих разрешению в процессе дальнейших исследований.
Всякая температура, превышающая максимальную температуру роста данного микроорганизма, является для него летальной. Вегетативные клетки бактерий, дрожжей и плесеней уничтожаются почти мгновенно под действием нагревания паром при 100°, не представляя, таким образом, особых затруднений при стерилизации консервов; однако споры некоторых видов бактерий чрезвычайно устойчивы против действия тепла, и для их уничтожения необходимы длительные сроки и высокие температуры нагревания. Скорость отмирания их является функцией взаимозависимых времени и температуры; при прочих равных условиях чем выше температура, тем быстрее происходит отмирание клеток. Поэтому летальный режим нагревания для данного вида микроорганизма, т. е. летальная точка, определяется температурой и сроком нагревания. Из дальнейшего изложения будет видно, что на режим стерилизации консервов, обеспечивающий полное отмирание данных микроорганизмов, оказывают сильное влияние многочисленные факторы, например химические и физические свойства среды, в которой присутствуют данные виды микроорганизмов, их количество, возраст и пр. Следовательно, для точного определения летальной точки недостаточно знать только температуру и срок нагревания.
Причина отмирания. Механизм отмирания бактерий под действием тепла еще недостаточно изучен. Некоторые исследователи считали, что теплостойкость бактерий связана со стойкостью ферментов; более поздними исследованиями установлена несостоятельность этой гипотезы, основывавшейся на существовании термостойких белковых ферментов, связанных, в частности, с бактериальными спорами; было выдвинуто предположение, что способность термофилов к росту при температурах выше точки коагуляции специфических белков зависит от термостойкости их ферментов. Такой вывод довольно сомнителен. Исследованиями, опубликованными в 1950 г., установлено, что ферменты активно развивающихся термофилов быстро инактивировались при их температурном оптимуме; однако, согласно другому сообщению, выделенные из термофильного микроорганизма системы дегидрогеназы яблочной кислоты и цитохрома в виде «красной фракции» выдерживали нагревание при 65°. Широко распространена точка зрения, заключающаяся в том, что отмирание происходит в результате коагуляции клеточных белков, и, действительно, имеется много данных в пользу того, что факторы, воздействующие на коагуляцию белков, оказывают сильное влияние на термостойкость бактерий. Например, кислая или щелочная реакция ускоряет коагуляцию белков и вызывает также снижение термостойкости бактерий. Исследования, проведенные при помощи электронного микроскопа, показали, что протоплазма молодой колонии бацилл приобретает крупнозернистую структуру под действием 10-минутного нагревания в воде при 55° или выше. В солевом растворе начало образования такой зернистости видно при 40°, а для полного завершения процесса требуется температура выше 50°. Каррен, цитируя сообщение об опытах по изучению действия жирных кислот в среде с прорастающими спорами на термостойкость Clostridium botulinum, приходит к заключению, что липоидные компоненты клеток содействуют термостойкости микроорганизма; при этом он указывает, что, если отмирание под действием влажного нагревания вызывается денатурацией некоторых важных белков (а это вполне возможно), то имеются основания для предположения, что липоидные компоненты спор оказывают защитное действие на пептидные связи против гидролиза или денатурации и могут продлить срок выживания.
Тепловая коагуляция альбумина, типичного представителя белков, колеблется в зависимости от содержания в нем воды, причем наиболее быстрая коагуляция происходит при высоком содержании воды, замедляясь при его снижении. Опубликованы данные, показывающие, что, несмотря на почти одинаковое содержание воды в вегетативных клетках и спорах, термостойкость спор разных видов бактерий зависит от содержания в них свободной воды; считают, что указанная разница, с которой вегетативные клетки и споры реагируют на нагревание, объясняется содержанием связанной воды в клетках, т. е. тем, что в клетках спор большее количество воды находится в связанном виде и поэтому не может принимать участия в коагуляции клеточных белков. Методика определения содержания связанной воды, или воды гидратации в белках, применявшаяся в вышеуказанных исследованиях, была подвергнута критике. Обзоры других теорий относительно причины высокой термостойкости бактериальных спор неоднократно опубликовывались в литературе. Автор одного обзора пришел к заключению, что наиболее вероятным объяснением может быть разное содержание белков в спорах и вегетативных клетках. В другом обзоре указывается, что споры обладают антигенными свойствами, которые не наблюдаются в вегетативных клетках, и что это обстоятельство говорит о существовании важных различий в химическом составе молекул вегетативных клеток и спор, обусловливающих чрезвычайно высокую теплоустойчивость последних.
Факторы, воздействующие на теплоустойчивость микроорганизмов
Порядок отмирания
На основании многочисленных данных можно заключить, что отмирание бактерий под действием нагревания может быть выражено логарифмической кривой, т. е. что в каждую последующую единицу времени отмирают равные количества клеток (в % от общего числа выживших). За последние годы опубликованы данные, показывающие, что порядок отмирания спор не следует логарифмической кривой; исследования проводились с применением пищевых сред и суспензий в буферном растворе. Учитывая затруднения, связанные с подсчетом числа выживших спор при исследовании порядка их отмирания под действием нагревания, необходимо доказать, что результаты, отклоняющиеся от результатов логарифмического порядка отмирания, статистически несовместимы с последними (если желательно считать их подтверждением отмирания нелогарифмического порядка). Статистический анализ данных по отмиранию микроорганизма РА 3679 под действием нагревания в нейтральном фосфатном буфере, был проведен американскими исследователями; анализ показал наличие исходной стадии быстрого отмирания, в течение которой гибнет более половины Присутствующих спор; второй стадии, в которой отмирание происходит в соответствии с логарифмической кривой, и третьей стадии, в которой скорость отмирания нескольких выживших спор снижается.
Несмотря на наличие противоречивых данных, имеются многочисленные факты, показывающие, что в большинстве случаев порядок отмирания бактерий носит логарифмический характер. Некоторые исследователи придерживаются мнения, что гибель микроорганизмов - результат случайного явления, например распада или инактиваций одной молекулы в клетке. Другие исследователи утверждают, что одной из причин логарифмического порядка отмирания бактерий может быть квантовый характер процесса нагревания; на этом основании они делают вывод, что возможность выживания одной споры в нескольких банках может быть отнесена за счет разнородного действия, оказываемого тепловой обработкой на банки. Выдвинуто предположение также о том, что порядок отмирания является результатом неоднородности отдельных клеток в суспензии. Показано наличие колебаний в удельном весе спор в суспензии и возможность фракционирования данной суспензии центрифугированием, причем летальные точки разных фракций различны. Если такое объяснение (т. е., что отмирание не является просто результатом случайных явлений) правильно, то вполне можно ожидать отклонения от логарифмического порядка в отмирании спор.
Кривая, построенная на основании логарифмов чисел выживших клеток в суспензии бактерий или спор, вычерченная в зависимости от времени нагревания при постоянной температуре, называется кривой выживания или кривой скорости отмирания микроорганизмов под действием тепла (рис. 8). При логарифмическом порядке отмирания получаем линейную кривую, угол наклона которой определяет время десятикратного снижения, обозначаемое обычно буквой D. Эта величина - время нагревания в минутах, требующееся для снижения количества живых микроорганизмов в суспензии бактерий до 7ю от исходного количества или же, выражаясь иначе, время в минутах, требующееся для прохождения кривой через один логарифмический цикл на шкале выживших особей.
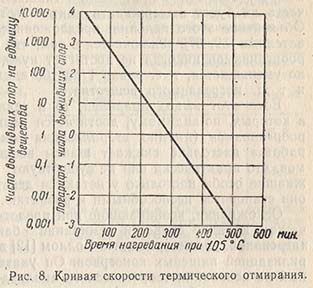
Откладывая значения D на логарифмической шкале в зависимости от соответствующих температур прямолинейной шкалы, получаем кривую «искусственных летальных точек»; угол наклона этой кривой, измеряемый необходимым для нагревания числом градусов Фаренгейта при прохождении кривой по одному логарифмическому циклу, равен значению 2 на кривой «термических летальных точек». Величину D можно получить, пользуясь кривой выживания, или вычислить ее на основании результатов нескольких пробных определений термической летальной точки, применяя методику, описанную американскими исследователями.
Рассмотрение типичной кривой термических летальных точек показывает, что, хотя теоретически количество выживших микроорганизмов может быть снижено до одного, какая-то часть их будет выдерживать длительный период нагревания. Объяснение этого явления, предложенное некоторыми исследователями, заключается в том, что число выживших микроорганизмов никогда не достигает нуля, а только прогрессивно уменьшается, т. е. равно 1 на 100 единиц, 1 на 1000 единиц и т. д. нагреваемого вещества.
Если этот вывод правилен, то в тех тепловых процессах, в которых, по-видимому, достигается полное уничтожение микроорганизмов (именно, стерилизация консервов), тепловая обработка настолько снижает шансы выживания микроорганизмов, что практически они не существуют, или же последние выжившие особи настолько угнетаются действием нагревания, что они становятся неспособными к развитию.
Осложнения, возникающие при представлении о логарифмическом порядке скорости отмирания бактерий под действием нагревания, рассматривались Боллом в связи с тепловой стерилизацией пищевых консервов. Он указывает, что «... сделанный на основании теоретического анализа вывод о том, что начальная микробиальная загрязненность пищевого продукта не может быть доведена до нуля в результате тепловой стерилизации, носил бы отрицательный характер, если бы отсутствовал практический опыт. За последние 15 лет, в течение которых были разработаны режимы стерилизации с использованием данных, основанных на термических летальных точках, в предположении, что таким образом обеспечивается полное уничтожение микроорганизмов, была доказана обоснованность этого метода с практической точки зрения. Во всяком случае доказано, что если даже в пищевом продукте остаются живые микроорганизмы, они находятся в столь угнетенном состоянии, что не могут принести вреда».
Влияние количества клеток
Из логарифмической кривой отмирания следует, что чем выше исходное количество клеток, тем больше потребуется времени для их полного уничтожения. Однако это замечание правильно только для клеток из идентичных суспензий. Термостойкость отдельной суспензии является ее индивидуальным свойством, редко совпадающим со свойствами других суспензий, даже если они были получены в аналогичных условиях. Однако несомненно для каждой данной суспензии, что при одних и тех же условиях нагревания термическая летальная точка будет выше, если увеличится концентрация клеток. На основании указанной зависимости термостойкости от количества клеток можно ожидать, что чем выше начальная загрязненность пищевого продукта, тем больше потребуется времени для его стерилизации. Промышленный опыт полностью подтверждает этот вывод. В качестве примера можно привести работу Камерона и др. с сахаром, обсемененным термофильными плоскокислыми спорами. Результаты их опытов подтвердили зависимость степени порчи от степени обсемененности продукта (табл. 4).
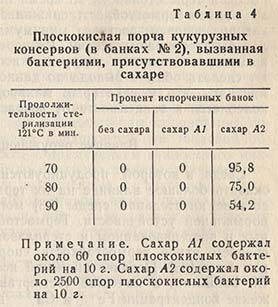
При тепловой стерилизации консервов большое значение имеет доведение исходного обсеменения продукта до минимума.
Резкое повышение степени обсемененности сырья или оборудования может привести к ненадежности режима стерилизации, даже вполне отвечающего своему назначению в течение ряда лет.
Влияние возраста микроорганизмов
Зависимость термостойкости микроорганизмов от их возраста изучалась многими исследователями с разными результатами. Согласно данным Эсти и Мейера, во влажной среде молодые споры Clostridium botulinum более термостойки, чем старые. Соммер утверждал, что споры Cl. botulinum достигали максимальной термостойкости по прошествии 4-8 суток выдержки в термостате. При изучении спорообразующих аэробов было установлено, что в процессе вызревания спор в течение годового периода термостойкость их повышалась, причем в первые три месяца значительных изменений не наблюдалось. Вильямс при исследовании различных видов спорообразующих аэробов и анаэробов не обнаружил никакой зависимости между возрастом культуры и ее термостойкостью в течение периода от 5 до 30 суток. Максимальная термостойкость спор штамма Clostridium sporogenes была достигнута в возрасте 7-14 суток. В этой связи изучались также неспорообразующие микроорганизмы. Было отмечено повышение устойчивости к теплу стрептококков во время начальной стадии логарифмического роста. Используя Escherichia coli в качестве тест-организма, исследователи установили наличие более высокой термостойкости во время начальной стационарной фазы, причем термостойкость снижалась, когда начиналась стадия воспроизведения, достигая минимума во время периода наиболее быстрого размножения. Разноречивые данные, полученные вышеперечисленными и другими исследователями, не позволяют сделать общего вывода по данному вопросу; однако можно признать, что у некоторых микроорганизмов термостойкость колеблется в зависимости от возраста культуры.
Влияние окружающей среды
Среда, в которой продуцируются опоры бактерий, может оказать большое влияние на их термостойкость. Изменения в составе питательной среды могут вызвать отклонения от нормальной устойчивости. Термостойкость спор, продуцированных в пастеризованном и стерилизованном мясе, превышала в среднем втрое термостойкость спор, продуцированных в сыром мясе. Позднее японский исследователь, используя Cl. botulinum в качестве тест-организма, установил, что снижение концентрации Fe++ и Са++ в среде, в которой выращивались споры, ниже определенной величины вызывало снижение термостойкости спор. В присутствии жирных кислот, в особенности кислот с длинными цепочками, наблюдалось повышение термостойкости спор.
Температура, при которой происходит рост микроорганизмов, также воздействует на их термостойкость; отмечено, что термостойкость спор увеличилась при повышении температуры выращивания последних до оптимальной величины. По данным вышеупомянутого японского исследователя, споры Cl. botulinum, выросшие при 37°, были более термостойкими, чем споры, развившиеся при 41, 28 или 24°. Другим исследователем установлено, что споры с максимальной температурой роста обладают максимальной термостойкостью.
Теплоустойчивость спор может колебаться в зависимости от физической природы среды, в которой они нагреваются. Исследуя вопросы, связанные с уничтожением термостойких спор в пищевых продуктах, Камерон и др. доказали наличие зависимости термостойкости спор от состояния продукта, т. е. от того, представляет ли он неоднородную массу или имеет пюреобразную форму.
Данные Вильямса и Рийда, подтвержденные данными более поздней работы, показали, что температура термостатной выдержки культур, выделенных из спор Cl. botulinum, подвергавшихся нагреванию, оказывает влияние на их термостойкость. Эти данные имеют практическое значение при бактериологических анализах консервов. Культуры, подвергавшиеся выдержке в термостате при 24, 27 и 31°, давали более высокий процент роста, чем параллельные культуры, выдерживавшиеся при 37°.
Состав среды, в которой помещаются микроорганизмы после нагревания, также может оказать значительное влияние на время их термического отмирания. Исследованиями установлено, что бактерии, подвергавшиеся нагреванию при сублетальной температуре, обнаруживали большую чувствительность в отношении условий роста по сравнению с бактериями, не подвергавшимися нагреванию. Кроме того, имеются указания, что чувствительность спор к ингибиторам прорастания повышается с усилением режима предварительной тепловой обработки спор. В работе с применением гнилостного микроорганизма РА 3679 было отмечено важное значение степени анаэробиоза; культуры, выращенные на печеночном бульоне с прослойками из тиогликоллатового агара, показали большую длительность выживания, чем развившиеся на простом агаре, применявшемся в качестве прослойки в питательной среде.
Предполагают, что в естественных средах, например в почве, споры могут обнаружить неодинаковую степень термостойкости по сравнению с искусственными средами. Одни исследователи утверждают, что опоры, образовавшиеся и созревшие в почве, более термостойки, чем споры, развившиеся в искусственных средах; было обнаружно, что термостойкость термофильных анаэробов становится значительно выше при включении почвы в питательную среду.
Однако в другой работе отмечается отсутствие разницы между термостойкостью спор анаэробных микроорганизмов, выделенных непосредственно из почвы или полученных путем посева. Единственным исключением была Clostridium sphenoides, найденная один раз; ее нативные споры обладали значительно более высокой термостойкостью, чем споры остальных культур. В литературе имеются указания, что для стерилизации почвы с естественным содержанием спор требуется в 6-8 раз более длительное нагревание, чем для уничтожения того же количества спор в культуре, выделенной из этой почвы. На основании этих данных был сделан вывод, что более высокая термостойкость спор в естественной почве возникает вследствие какого-то физико-химического действия почвы, а не вследствие различий между спорами из естественной почвы и выращенными на искусственных средах.
Учитывая все имеющиеся данные, можно, по-видимому, заключить, что результаты лабораторных исследований по теплоустойчивости спор, выращенных на искусственных средах, нельзя считать полностью соответствующими условиям заводской практики. Камерон и др. изучали спорообразующий микроорганизм, вызвавший порчу консервированной свеклы, стерилизованной при 116° в течение 20 мин. На основании данных по прогреванию содержимого банок этот режим должен быть достаточным для гибели спор, выдерживающих примерно 10-минутное нагревание при указанной температуре. В лабораторных условиях, однако, после 5-минутного нагревания при 110° выживших спор не было. Исследователи заключили, что исходная термостойкость спор должна была быть значительно выше, чем, показанная лабораторными опытами, и что снижение термостойкости, по-видимому, произошло за счет методики опыта, применявшейся в данном случае.
Влияние величины pH
Концентрация водородных ионов в питательной среде может оказать большое влияние на термостойкость микроорганизмов. Для большинства спорообразующих бактерий максимальная устойчивость обычно находится в нейтральной зоне. Эсти и Мейер установили максимальную стойкость Clostridium botulinum при значениях pH от 6,3 до 6,9. Для спор Bacillus subtilis обнаружена максимальная термостойкость при pH 6,8-7,6. Дрожжи имели несколько более высокую термостойкость при pH 3,8-4,0. Однако за одним только исключением, штаммы дрожжей, изучавшихся Таннером и др., имели более высокую термостойкость при pH 6,8 по сравнению с pH 3,8. Позднее, с использованием Saccharomyces sp. в качестве тест-организма, было установлено, что он более устойчив к нагреванию ;в буферной среде ,с pH 5,6, чем при pH 4,0. Максимум термостойкости аскоспор Byssochlamys fulva находится примерно при pH 5,0; кроме того, стойкость при pH 3,0 значительно выше, чем при pH 7,0. Опытами автора, проведенными в 1940 г., установлено, что неспорообразующие бактерии обнаруживают максимальную стойкость к теплу и к дезинфицирующему действию спирта и наибольшую стойкость во взвешенном состоянии при одном и том же значении pH. На этом основании был сделан вывод, что при этом значении pH гидратация белков бактерий также достигает максимума и поэтому микроорганизмы менее восприимчивы к действию вредных для них агентов.
Повышение концентрации водородных ионов обычно вызывает соответствующее снижение термостойкости. В отношении консервированных пищевых продуктов влияние умеренных изменений величины pH на термостойкость спор, по-видимому, не всегда одинаково. Эсти и Мейер утверждали, что при pH выше 5,0 на время стерилизации консервов скорее влияют не концентрация водородных ионов, а другие факторы. В отношении консервированных морских продуктов никакой связи между величиной pH и термостойкостью спор Clostridium botulinum в пределах значений pH 5,2-6,8 не найдено. Однако при pH 4,9 наблюдалось резкое снижение термостойкости. Более поздними исследованиями [59] установлено, что изменение pH в пределах узкого интервала значений, обычно встречающихся в консервах (5,95-7,25), в сливках, в которых были подвергнуты нагреванию споры Bacillus subtilis, вызывало колеблющиеся результаты. В литературе имеются указания, что чем ниже значение pH в пределах 4,5-9, тем слабее должно быть нагревание для предупреждения порчи овощных консервов разных видов. При значениях pH около 5,5 была отмечена ясно выраженная необходимость в повышении режима тепловой обработки, обеспечивающего «коммерческую» стерильность консервов. При значениях pH в пределах 6-9 требуется лишь незначительное повышение температуры нагревания с увеличением pH.
В отчете, опубликованном в 1954 г., отмечается, что в пищевых средах изменения pH могут иметь не только прямое действие на микроорганизмы, но могут вызвать изменения в степени диссоциации веществ или в окислительно-восстановительном потенциале, которые в свою очередь могут оказать косвенное влияние либо на выживание, либо на способность выживших микроорганизмов к росту.
Андерсон и др., исследуя выживание В. thermoacidurans (В. coagulans) в томатном соке, установили наличие снижения температуры отмирания после добавления разных количеств уксусной, молочной и лимонной кислот. Базируясь на процентном содержании добавляемых кислот, последние можно расположить в следующем порядке по степени снижения летальной точки: молочная > лимонная > уксусная; базируясь на величине pH, придется принять следующий порядок: уксусная > молочная > лимонная. Добавочное действие кислотности на микроорганизмы, разрушаемые нагреванием, используется при стерилизации некоторых овощных и других некислотных консервов, режим стерилизации которых без добавления указанных кислот будет слишком резок и нанесет ущерб их органолептическим качествам. В таких консервах подкисляют заливку, в результате чего снижается термостойкость загрязняющих их микроорганизмов. Этот способ широко применяется в Америке при производстве консервов из артишоков.
Действие соли
Многочисленные исследования, проведенные по изучению действия соли на термостойкость микроорганизмов, не дали полностью согласованных результатов. В 1921 г. было установлено наличие значительного снижения термостойкости спор Clostridium botulinum в присутствии 3% соли; при более высоких концентрациях наблюдалось резкое снижение термостойкости. Позднее было найдено, что термостойкость этого микроорганизма повышалась в присутствии 1-2% соли, а при наличии свыше 8% соли стойкость снижалась. Термостойкость Clostridium welchii повышалась в присутствии 3% соли; при 10%-ном содержании ее было отмечено ясно выраженное снижение термостойкости. Опубликованы также данные, показывающие наличие снижения термостойкости спор Cl. botulinum в фосфатной буферной среде с pH 7,0 в присутствии 3,5% соли при температурах ниже 110°; однако в пределах ряда температур 110-112,7° никакого заметного действия обнаружено не было.
По данным других исследователей, соль в концентрации до 4% оказывала защитное действие в отношении спор нескольких видов микроорганизмов, подвергаемых нагреванию в заливочном растворе консервированного горошка. Андерсон и др. установили наличие прогрессивного снижения летальной точки для Bacillus thermoacidurans в присутствии 1, 2, 4 и 8% соли в томатном соке, хотя частичное действие могло относиться за счет незначительного снижения величины pH в результате добавления соли. Согласно данным ряда американских исследователей, добавление соли и других консервантов (нитрат натрия и нитрит натрия) в концентрациях, обычных для обработанных ими продуктов (солонины), не оказывает заметного влияния на режим стерилизации, требующийся для консервированной солонины и подобных продуктов.
Андерсон и др., проводившие исследования с дрожжами в подкисленной среде, нашли, что соль в концентрациях 7,5 и 10% обладала слабым защитным действием, но при 15% термическая летальная точка несколько снижалась. В литературе имеются указания на защитное действие соли в отношении термостойкости микрококков при нагревании.
По-видимому, действие соли колеблется в зависимости от ее концентрации, от среды и вида тест-организма. Частично это действие может быть вызвано побочными эффектами, например изменением величины pH или степени диссоциации прочих растворенных веществ.
Действие сахара, белков и других веществ
Сахар, глицерин и прочие нетоксические вещества оказывают защитное действие на микроорганизмы при нагревании. В присутствии таких веществ, взятых в достаточной концентрации, их защитное действие в некоторых случаях является настолько ясно выраженным, что получает большое значение в производстве консервов. Дрожжи быстрее погибают при 100° в дистиллированной воде, чем в сиропе. Термическая летальная точка в сиропе с концентрацией 24° Боме соответствовала 5 мин., а в сиропе при 36° Боме - 28 мин. В присутствии 10, 25 и 50% сахара термостойкость плесеней повышалась. Автор нашел, что срок выживания Escherichia coli при 70° при добавлении 10% сахарозы увеличился с 4 до 6 мин.; в 30%-ном растворе сахарозы указанные микроорганизмы выдерживали нагревание в течение 30 мин.
Действие различных концентраций сахара на термостойкость некоторых микроорганизмов, вызывающих порчу консервов, приводится в табл. 5, взятой из отчета Брауна и др.
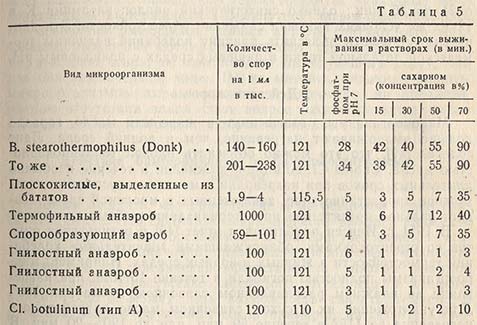
Проведенные указанными авторами исследования по изучению причин порчи консервированных бататов ясно показали практическое значение защитного действия сахара на микроорганизмы. Они установили, что порча была вызвана добавлением сухого сахара вместо обычного сахарного сиропа. Сахар не растворился и не был равномерно распределен в продукте, как ожидали, но образовал толстый слой густого сахарного сиропа, который защищал микроорганизмы-возбудители порчи против действия тепла при нормальной стерилизации.
Опыты с Cl. botulinum показали увеличение термостойкости этого микроорганизма при повышении концентрации сахарозы, причем было отмечено некоторое ее защитное действие в коцентрациях ниже 12,5%. Опубликованы также сообщения о повышении термостойкости Bacillus thermoacidurans в томатном соке, к которому были добавлены сахароза или декстроза.
Помимо сахара, защитное действие на микроорганизмы против нагревания оказывают также белки, например желатин и сывороточные белки. Отмечено также и повышение летальной термической точки спор Clostridium tetani при pH 7,0 и повышенном содержании органического вещества (пептона).
При изучении влияния некоторых витаминов на термостойкость спор анаэробов было отмечено, что добавление 0,01% аскорбиновой кислоты к нейтральному фосфатному буферному раствору вызывало повышение термической летальной точки видов Clostridium, при котором срок нагревания увеличивался с 24 до 36 мин.; однако синтетический аналог витамина К в таких же условиях вызывал снижение термостойкости. Эти данные могут отчасти объяснить причину колебаний величины термической летальной точки в пищевых средах с одинаковым pH.
Действие жиров
Микроорганизмы, взвешенные в жире или масле, гораздо дольше выдерживают нагревание, чем в водной среде. Ланг полагал, что это можно объяснить плохой теплопроводностью масла. Споры Cl. botulinum выживали в течение необычно длительных сроков при нагревании в масляных суспензиях; возможное прорастание спор, заключенных в масляной среде, создает сильное препятствие при стерилизации рыбных консервов.
В 1954 г. Йенсен опубликовал отчет о результатах опытов со стрептококками, обнаружившими значительные колебания термостойкости в жирных и водных средах. Например, микроорганизмы полностью погибали в течение 15 мин. при нагревании во влажном расплавленном сливочном масле при 100°. Для уничтожения их в сухом сливочном масле потребовалось нагревание до 115° в течение 50 мин. или до 120° - 20 мин. В молоке микроорганизмы отмирали при нагревании до 61,7° в течение 30 мин.
Рогачева установила, что термостойкость неспорообразующих бактерий не увеличивалась при нагревании в жире или в масле. Термостойкость спор групп Subtilis-mesentericus увеличивалась, но только при применении обезвоженных жиров или масел. В том же году Николс сообщил, что повышение содержания молочного жира в молоке или сливках не увеличивало термостойкости спор Bacillus subtilis.
Отмечено, что дрожжи Saccharomyces sp. обладали более высокой термостойкостью в масляной среде, чем в буферном растворе с одинаковым значением pH; для стерилизации французского салатного соуса потребовалась температура выше применяемой для уничтожения этих дрожжей в водной среде.
Особый интерес с точки зрения производства консервов представляет работа Камерона и др. по изучению защитного действия жира на кокки. В этих опытах применялись культуры микрококков, выделенных из стерилизованных жирных продуктов. Влажные кокки, подвергавшиеся нагреванию в бульоне, отмирали через 30-45 мин. при 55°, между тем как кокки, подвергавшиеся сначала сухому нагреванию, а затем нагреванию в бульоне, обладали несколько более высокой термостойкостью и отмирали через 45-60 мин. при 55°. Для уничтожения сухих кокков во влажном жире необходимо было нагревать их 15-20 мин. при 100°. В сухом жире наблюдалось ясно выраженное повышение термостойкости; указанные микроорганизмы выдерживали 135-минутное нагревание при 100°. На основании имеющихся данных авторы считают, что микрококки, выжившие после стерилизации благодаря защитному действию жира, не способны к последующему росту, так как они заключены в жире. По их мнению, механизм защиты микроорганизмов жиром основан, по-видимому, на локализованном отсутствии влаги. Этот вопрос рассматривается также в главе по микробиологии доброкачественных консервов.
Действие бактерицидных и бактериостатических веществ
Можно ожидать, что присутствие в нагреваемой среде бактерицидных и бактериостатических веществ вызовет снижение термостойкости микроорганизмов. В литературе имеются указания о значении применения небольших количеств гермицидов для увеличения эффективности низких режимов стерилизации. Присутствие сернистого ангидрида в концентрации 0,0022% может дать возможность снизить температуру стерилизации фруктовых соков примерно на 5°. Отмечено также, что очень малые концентрации сернистого ангидрида, которые могут быть введены в консервированные фрукты с сахаром, вызывают резкое снижение термостойкости аскоспор Byssochlamys fulva. Например, время, необходимое для уничтожения около 99% указанных микроорганизмов при 85° (pH 3,0), было снижено с 40 до 14 мин. добавлением 0,0002% сернистого ангидрида. Более поздними исследованиями было установлено, что концентрация сернистого ангидрида для достижения данного летального действия зависит от pH нагреваемой среды, причем концентрация должна увеличиваться при повышении pH.
При изучении действия эфирных масел некоторых специй на термостойкость микроорганизмов было установлено, что добавление аллилизотиоцианата (летучее эфирное масло горчицы) в концентрации 0,001% в буферные растворы или в яблочный и виноградный соки вызывало резкое снижение термостойкости Aspergillus niger и Saccharomyces ellipsoideus. Действие это было слабее выражено в отношении Bacillus thermoacidurans.
Эфирные масла чеснока или лука менее эффективны, чем аллилизотиоцианат. Было также отмечено, что в пределах практического применения эфирные масла гвоздики, корицы и препарата чеснока с укропом оказали незначительное действие на споры Bacillus thermoacidurans, суспендированные в томатном соке. Эфирные масла корицы и чеснока были эффективными в отношении снижения термостойкости дрожжей, вызывающих порчу маринованных огурцов; опыты проводились в подкисленном рассоле. Добавление препарата, состоявшего из эфирных масел чеснока и укропа в приемлемой для вкуса концентрации (0,0258%), и эфирного масла горчицы в концентрации 0,002% значительно снизило термическую летальную точку тест-организма дрожжей, суспендированного в подкисленном рассоле. На основании результатов этих опытов можно заключить, что эфирные масла некоторых специй могут оказать заметное действие на дрожжевые клетки, но это действие значительно слабее в отношении спор бактерий.
Неочищенные экстракты многих пищевых растений тормозят рост микроорганизмов. Некоторые экстракты при применении их в комбинации с пониженным режимом тепловой обработки снижают термостойкость бактерий плоскокислой порчи. Оба эти действия снижают срок выживания микроорганизмов в стерилизованных консервах и могут в некоторой степени объяснить колебания термостойкости данного микроорганизма в различных пищевых средах.
Антибиотики
Андерсон и Митченер были первыми, установившими (в 1950 г.) возможность использования антибиотиков (субтилина) в сочетании с пониженным режимом тепловой обработки для стерилизации консервов. Этот способ основан на уничтожении при слабом нагревании естественных ферментов и неспорообразующих бактерий, дрожжей и грибков и на задержании прорастания спор антибиотиком.
Исследователи приводят результаты опытов с овощными консервами, стерилизованными комбинированной обработкой - добавлением субтилина в концентрации 0,0005-0,002% и нагреванием при 100° в течение 5-10 мин. Преимущества такого способа стерилизации совершенно очевидны; не только устраняется отрицательное действие избыточного нагревания на органолептические качества консервов, но значительно сокращаются производственные издержки. Однако позднейшими исследованиями результаты этих опытов подтверждены не были. Камерон и др. указали, что опыты с антибиотиками не выдержали основного требования стерилизации - уничтожения спор Cl. botulinum. Они отметили также, что субтилин, с которым было проведено большинство опытов, оказался неэффективным против сапрофитных микроорганизмов-возбудителей порчи пищевых продуктов.
Тот факт, что субтилин, взятый в относительно высокой концентрации, в сочетании с пониженным режимом тепловой обработки, оказался недостаточно активным против Cl. botulinum, подтвержден и другими исследованиями; такие же результаты были получены с другими антибиотиками (грамицидин, метилограмицидин, бацитрацин и стрептомицин).
Опубликованы также данные о том, что субтилин в сочетании со слабым нагреванием не уничтожал спор ряда термофильных микроорганизмов, выделенных из испорченных консервов. Адамс и др., работавшие с говяжьим фаршем и спорами гнилостного анаэроба, установили, что высокие концентрации субтилина в комбинации со слабой тепловой обработкой не давали полного торможения роста спор в говядине; для этого требовалось применение сильной тепловой обработки и высоких концентраций субтилина.
Позднее была исследована термостойкость спор Clostridium botulinum и гнилостного анаэроба РА 3679 в пюре из горошка. В присутствии субтилина в концентрации 0,0014% термостойкость РА 3679 составляла 47% по сравнению с термостойкостью в пюре из горошка без добавления субтилина. В той же суспендированной среде споры Clostridium botulinum обладали термостойкостью, составлявшей 63% от нормальной. В этих опытах применялись температуры в пределах 115,5-132,2°.
Из сказанного выше следует, что при существующем в настоящее время положении стерилизация пищевых консервов слабой тепловой обработкой, комбинированной с антибиотиками, не является надежным способом. Многочисленные данные показывают, что некоторые антибиотики снижают термостойкость спорообразующих микроорганизмов, обычно присутствующих в консервах; однако требуется проведение дальнейшей значительно расширенной и углубленной исследовательской работы, включающей изучение возможных физиологических эффектов антибиотиков, прежде чем последние можно будет безопасно применять в производстве консервов.
Действие сублетальной тепловой обработки
Состояние покоя
Замедленное прорастание или состояние покоя прогретых спор наблюдалось и изучалось многими исследователями как вопрос, которому придавалось особое значение в микробиологии консервов. Споры, подвергавшиеся сублетальной тепловой обработке, иногда даже в благоприятной среде остаются в состоянии покоя в течение длительного времени. Отмечено, что прогретые споры Cl. botulinum могут оставаться в таком состоянии в течение шести лет и более. Эсти и Мейер сообщали, что прогретые суспензии спор Cl. botulinum оставались в состоянии покоя в течение 378 суток при 36-37°. Эти случаи исключительны; однако периоды покоя, длящиеся неделями, наблюдаются часто при производстве бактериологических анализов консервов.
Причина состояния покоя - все еще спорный вопрос. Предполагалось, что оно обусловлено селективным действием нагревания на клетки с толстыми, относительно непроницаемыми стенками. Считается вероятным и то, что в результате тепловой обработки клеток последние подвергаются какому-то повреждению, мешающему их нормальному развитию.
Весьма трудно себе представить, чтобы фактор окружающей среды не влиял на состояние покоя микроорганизмов. Рядом исследователей установлено, что состояние покоя прогретых микроорганизмов может быть нарушено или ослаблено наличием благоприятной для роста среды. При исследовании процесса прорастания спор Cl. botulinum было отмечено, что добавление 0,1% растворимого крахмала в среду оказывало сильное действие на процесс прорастания. Крахмал действует в основном путем поглощения небольших количеств присутствующих в среде веществ, подавляющих прорастание спор. Те же исследователи установили, что для прорастания спор Cl. botulinum в синтетической среде необходимо присутствие небольших количеств углекислоты; при отсутствии углекислого газа значительно задерживается прорастание в сложной органической среде. На другие анаэробы углекислый газ не действовал; однако опубликовано сообщение об увеличении количества микроорганизмов РА 3679 при добавлении в среду бикарбоната натрия. Андерсон и др. подтвердили необходимость включения бикарбоната натрия в среды, используемые для подсчета спор Cl. botulinum. Значение этих данных в связи с выделением бактерий из продуктов, подвергавшихся нагреванию, совершенно очевидно.
Стимулирование роста нагреванием и активация
При изучении сублетального нагревания спор, помимо состояния покоя, наблюдались также и другие явления. Исследованиями установлено наличие большего увеличения термостойкости, обнаруженное у штаммов Cl. botulinum и у одного гнилостного анаэроба при выдерживании культур при 24, 27 и 31°, по сравнению с термостатной выдержкой при 37°, несмотря на то, что температурный оптимум роста этих микроорганизмов находился в пределах 35-37°. Опубликованы также данные, показывающие, что относительно слабое нагревание вызывает более быстрое прорастание спор мезофильных аэробов; в этих работах изучалась группа термостойких аэробов, выделенных из испорченного консервированного сгущенного молока без сахара; в результате было установлено, что сублетальное нагревание спор снижает минимальную температуру, при которой может иметь место рост. Такое прорастание спор при относительно низкой температуре, называемое «тепловой активацией», наблюдается в тех случаях, когда нагревание является слабым и не дает летального эффекта (10-30 мин. при 95°). Об активации спор слабым нагреванием сообщали Стэмбо, Мэрфи и др., работавшие с гнилостным анаэробом РА 3679. В отношении консервированных пищевых продуктов очень важное значение имеет тот факт, что тепловая активация может быть причиной прорастания термофильных спор в пределах нормального для хранения консервов интервала температур.
Методы определения теплоустойчивости микроорганизмов
Наличие большого числа факторов, влияющих на теплоустойчивость микроорганизмов, вызвало необходимость в разработке точно стандартизированной методики, обеспечивающей получение надежных данных; для этого необходимо проведение ряда повторных проб, и только после этого можно будет пользоваться полученными данными по термостойкости определенного вида микроорганизмов при разработке режимов стерилизации. Ниже перечислены некоторые важные факторы, которые следует учитывать при установлении летальной термической точки, с использованием одних и тех же микроорганизмов в одних и тех же средах:
1. Колебания в термостойкости отдельных спор в данной суспензии.
2. Изменения в термостойкости спор в запасной суспензии во время хранения.
3. Потеря части культуральной суспензии при перенесении из нагретой банки в питательную среду.
4. Изменения в различных образцах одной и той же питательной среды, влияющие на пригодность их для выращивания спор.
5. Возможное отсутствие прорастания нескольких выживших спор во время термостатной выдержки. Для многих микроорганизмов необходимо определенное количество спор для получения показательного роста или даже прорастания.
Фактором, значение которого прогрессивно увеличивается в связи с промышленным применением асептического метода консервирования и с необходимостью наличия данных о термической летальной точке при температурах выше 121,1°, является летальное действие всего периода, в течение которого температура тест-организма доводится до температуры стерилизации. Именно по этой причине последние разработанные методы определения летальной термической точки (или скорости отмирания микроорганизма) основаны на мгновенном нагревании и охлаждении образцов.
В тех случаях, когда нагревание проводится в фосфатном буфере или какой-либо другой непищевой среде, необходимо произвести пересев культуры в питательную среду. За последнее время наблюдается стремление к получению этих данных на спорах, суспендированных и нагреваемых в пищевом продукте, когда определение процента выживания может проводиться путем пересева или термостатной выдержки продукта. Полученные результаты могут колебаться в зависимости от того, применялся ли пересев или выдержка продукта в термостате. Это, несомненно, объясняется многими факторами, наиболее важными из которых являются влияние величины pH, присутствие тормозящих агентов и необходимые для выживших микроорганизмов условия питания.
Ниже приводится краткое описание методик определения термостойкости микроорганизмов, применявшихся при решении вопросов, связанных со стерилизацией консервов.
Метод Бигелоу и Эсти
Метод известен под названием «метод одной трубочки» (пробирки); он заключается в нагревании определенного количества суспензии микроорганизмов или спор в небольших стеклянных трубочках (диаметр в свету 7 мм, толщина стенок 1 мм, длина 250 мм). После запаивания на пламени трубочки погружаются в масляную баню с постоянной температурой; через короткие промежутки времени удаляют по одной трубочке, быстро охлаждают ее и делают посев (выливают содержимое) в соответствующую среду. По этому методу термическим временем отмирания считается период между временем нагревания последней трубочки, давшей рост на субкультуре, и временем нагревания первой трубочки, не показавшей роста. Недостатком этого метода является наличие «скачков», под которыми подразумеваются трубочки, не давшие роста на субкультуре, хотя они нагревались в течение более короткого срока, чем трубочки с сохранившимися живыми микроорганизмами.
Метод Эсти и Вильямса
Этот метод был разработан с целью устранения скачков предыдущего метода. Вместо удаления с нагревательной бани по одной трубочке извлекают по 25-30 трубочек четыре раза через довольно большие интервалы. Трубочки быстро охлаждаются, и их содержимое переносят в соответствующую среду. Для каждого периода нагревания на полулогарифмической бумаге отмечается процент трубочек, давших рост; на основании полученной прямой линии можно найти путем экстраполяции термическую летальную точку для данного микроорганизма при данной температуре.
Эсти, рассматривая относительные достоинства и недостатки методов с применением одной трубочки и большого количества трубочек, приводит типичный пример, когда метод одной трубочки показал полное уничтожение опор через 20 мин., между тем как при применении метода со многими трубочками 39,4% трубочек дали рост при наличии тех же условий. Он отмечает, что метод с применением одной трубочки полезен при указании интервала, в пределах которого достигается почти полное уничтожение спор, за исключением нескольких случайных. Метод с применением многих трубочек наиболее пригоден при подсчетах, связанных с разработкой режимов стерилизации.
Метод Вильямса, Меррила и Камерона
Указанные исследователи разработали аппарат, дающий возможность нагревать суспензии под давлением и отбирать образец без; нарушения условий нагревания. Этот аппарат, представляющий собой автоклав в миниатюре, оборудован пропеллером для перемешивания суспензии и специальными клапанами для отбора образцов; аппарат сконструирован для определения скорости уничтожения спор. По утверждению изобретателей, аппарат этот более удобен и дает более точные результаты, чем метод Эсти и Вильямса. Несколько модифицированная конструкция этого аппарата была изготовлена Научно-исследовательским отделом английской компании Метал Бокс Компани (рис. 9).

Метод Тоунсенда, Эсти и Базелта
Для определения термической летальной точки сырых пищевых продуктов указанные исследователи применяли специальные банки 63,5 мм в диаметре и 9,5 мм высотой. Эти банки, содержавшие сырой продукт и тест-организмы, закатывали под вакуумом, нагревали в небольших автоклавах специальной конструкции, быстро охлаждали и подвергали термостатной выдержке. Рост тест-организма устанавливали по вспучиванию банок, вызванному газообразованием.
Позднее в такие банки после нагревания добавляли концентрированную среду, прокалывая банку и вводя среду при помощи шприца и иглы; после запаивания отверстия кусочком припоя банки подвергали термостатной выдержке обычным порядком. В большинстве случаев при применении дополнительных сред из пищевого продукта наблюдались более длительные сроки выживания, чем при непосредственной выдержке в термостате.
Метод Стэмбо, Гросса и Винтона
При изучении тепловой стерилизации мясных консервов указанные авторы нагревали инокулированный мясной фарш в стеклянных трубочках размером 10x75 мм. Трубочки наполняли продуктом при помощи тавотного шприца с иглой, применяющейся для вливания в артерию; наполненные трубочки запаивались в пламени и нагревались на бане с лярдом. Авторы перечисляют следующие достоинства этого метода: 1) возможность широкого варьирования условий нагревания и условий обработки мяса, возможность проведения повторных опытов по каждому варианту без большой затраты времени и места; 2) легкость введения мяса в стеклянные трубочки механическим путем; 3) возможность быстрого перемещения содержимого трубочек после стерилизации в среды, благоприятные для роста тест-организмов, с очень небольшим риском загрязнения; 4) возможность выдержки в термостате невскрытых трубочек с мясом после стерилизации; 5) легкость наблюдения прогрессивной порчи стерилизованного мяса в стеклянных трубочках и период термостатной выдержки.
Метод Стэмбо
Прибор терморезистометр, описание которого было опубликовано в печати указанным исследователем в 1948 г., предназначался для изучения термостойкости бактерий в интервале температур от 115,6 до 132,2°. Прибор состоит из трех камер, присоединенных к резервуару с паром и сконструированных таким образом, что каждая камера может работать под давлением или с удалением отработанного пара, независимо от остальных двух камер. Инокулированные образцы, разлитые в небольшие металлические чашечки, диаметром около 7 мм и высотой 1,5 мм, помещают в алюминиевые лоточки. Образцы проходят по очереди через три, предварительно стерилизованные камеры. В первой камере чашечки наполняются; во вторую, являющуюся контрактной камерой, подается пар при рабочей температуре для создания давления; в третьей камере образцы, охлажденные выпариванием, поступают в пробирки с культуральной средой. Время стерилизации в центральной камере точно измеряется при помощи электрических часов, регулируемых микровыключателями, которые срабатывают, когда образец входит в камеру и когда выключается давление пара. Преимущества этого способа, по утверждению изобретателя, заключаются в следующем: 1) достижении более высокой точности, в особенности в интервале высоких температур, вследствие быстрого нагревания и охлаждения образцов; 2) относительной простоте работы; 3) снижении опасности загрязнения и др.
Метод Шмидта
По этому методу тест-суспензию нагревают в небольших стеклянных трубочках, укупориваемых пробками из хлопка-сырца, которые позволяют добавлять субкультуральную среду непосредственно в трубочки. Разбрызгивание жидкой среды и срыв пробок предотвращаются системой охлаждения под давлением, присоединенной к миниатюрному автоклаву, применяемому для операций нагревания и охлаждения. Термопара, установленная в одной из трубочек, позволяет более точно измерить отставание температуры, допуская возможность использования аппарата при высоких температурах, порядка 121,1-132,2°.
Метод Пфлуга и Эсселена
Эти исследователи пользовались прибором, сходным по конструкции с терморезистометром. Образцы в металлических чашечках вводили в паровую камеру и удаляли из нее при помощи несущего патрона. Наполнение чашечек и пересев производились в другой камере; для охлаждения образец смешивали со средой, в которую производился пересев, а не выпаривали его, как в терморезистометре. Для предотвращения кипения жидкости после выхода из камеры, работающей под давлением, и перед автоматическим пересевом применялся стерильный сжатый воздух.
Метод Штерна и Проктора
По этому методу суспензию спор помещают в запечатанные капиллярные трубки с внутренним диаметром 0,8 мм, наружным диаметром 1,5 мм и длиной 76,2 мм. Трубки, удерживаемые на месте в лотке-носителе, автоматически погружаются в горячую баню, и в этот момент начинает работать электрический автоматический регулятор, времени. В конце заданного срока погружения свободно падающая свинцовая гиря весом в 2,3 кг начинает вращать барабан, который переносит лоток-носитель, проходящий по полукругу, из горячей бани в холодную; на это требуется 0,37-0,40 сек. Прибор применялся при изучении термостойкости микроорганизмов при температурах от 100 до 140° с использованием поправочных коэффициентов, выведенных на основании прогревания бесконечного цилиндрического тела. Преимуществом этого метода является возможность обработки 10 и более образцов одновременно, облегчающая статистический анализ полученных данных.