Лизин
Лизин (а, е-диаминокапроновая кислота) известен в виде двух оптически активных D- и L-форм и рацемической DL-формы. Эмпирическая формула C6H12N2O2. Молекулярная масса 146,19. Лизин хорошо растворим в воде, кислотах, основаниях; трудно растворим в спирте и нерастворим в эфире. Аминокислота при температуре 224-225 °С разлагается. Кристаллизуется лизин в виде бесцветных игл или гексагональных пластинок.
Установлено, что в организме лизин определяет не только биологическую ценность белка. Аминокислота выполняет много и других биохимических функций - она способствует секреции пищеварительных ферментов и транспорту кальция в клетки, улучшает общий азотный баланс в организме. Применение лизина в хлебопекарной промышленности повышает биологическую ценность и улучшает качество изделий. От добавления лизина в рационы животных (0,1-0,4%) значительно увеличивается коэффициент использования белка, и тем самым снижается расход кормов на единицу продукции.
Биосинтез лизина
Он осуществляется при помощи ауксотрофных мутантов микроорганизмов рода Micrococcus, Brevibacterium, Corynebacterium и др. Питательные среды, используемые для выращивания микроорганизмов и биосинтеза аминокислоты, содержат в качестве источника углеводов свекловичную мелассу, кукурузный экстракт или белковые гидролизаты - источники аминокислот. Источниками азота могут служить соли аммония, мочевина. При биосинтезе лизина важную роль играет концентрация факторов роста в среде - биотина и необходимых аминокислот - метионина, гомосерина, треонина. Для роста и биосинтеза лизина культурой Brevibacterium sp. 22 оптимальной считается следующая концентрация на 1 л питательной среды: метионина 200 мг, треонина 800 мг, биотина 15-20 мкг. При уменьшении концентрации биотина (на 1 л) до 1-4 мкг культура Brevibacterium sp. 22 синтезирует глутаминовую кислоту, при увеличении до 2,5 мг образуется молочная кислота - явление, известное как механизм обратного действия.
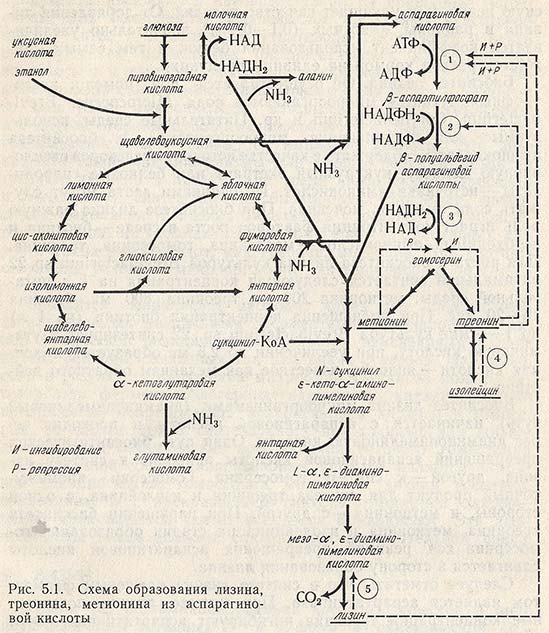
Биосинтез лизина микроорганизмами (диаминопимелиновый путь) начинается с аспарагиновой кислоты и проходит через диаминопимелиновую кислоту. Один путь биосинтетических превращений аспарагиновой кислоты приводит к синтезу лизина, другой - к синтезу гомосерина. Гомосерин - промежуточный продукт для синтеза треонина и изолейцина, с одной стороны, и метионина - с другой. При нарушении биосинтеза треонина, метионина и изолейцина на стадии образования гомосерина ход реакций превращения аспарагиновой кислоты сдвигается в сторону образования лизина.
Следует отметить, что в синтезе лизина ключевым ферментом является аспартаткиназа. При синтезе лизина повышенные концентрации треонина ингибируют аспартаткиназу. Этот эффект усиливает присутствие лизина. Треонин у бактерий (Е. coli, Micrococcus glutamicus) ингибирует дегидрогеназу полуальдегида аспарагиновой кислоты и гомосериндегидрогеназу. Метионин по отношению к гомосериндегидрогеназе является репрессором. Треониндегидрогеназу ингибирует изолейцин. Таким образом, продукты обмена веществ, угнетающие различные ферменты и участвующие в синтезе лизина, должны быть выведены из реакции. Именно поэтому ауксотрофные микроорганизмы являются наиболее удобными для производства. Например, культура, лишенная активности гомосериндегидрогеназы, обеспечивает достаточно высокие выходы лизина.
На рис. 5.1 приведена схема биосинтеза лизина в бактериальных клетках, на которой показаны промежуточные метаболиты, синтезируемые по общему с лизином пути из аспарагиновой кислоты.
Технологические схемы производства L-лизина. Технология получения кристаллического L-лизина, разработанная Всесоюзным научно-исследовательским институтом генетики и селекции промышленных микроорганизмов, состоит из двух основных стадий: культивирования продуцента и выделения конечного продукта. На стадии культивирования производится двухступенчатое выращивание посевной культуры Micrococcus glutamicus в инокуляторах и посевных аппаратах. Предварительно выраженной культурой засевается ферментатор, в котором в стерильных аэробных условиях при непрерывном перемешивании и термостатировании (30-33 °С) осуществляется биосинтез. При ферментации в течение 50-70 ч при pH 7,4 концентрация лизина в растворе достигает 40 г/л.
Полученная после биосинтеза культуральная жидкость направляется на осветление, в результате отделяется биомасса. Осадок отделяют фильтрацией, сушат, размалывают и используют как кормовой продукт, содержащий около 40 % белковых веществ.
Очищенный раствор с pH 7,0 пропускают через катионит марки КУ-2 или КБ-4П-2 в NH4+-форме. Каждые 100 г катионита сорбируют 6-8 г лизина. После промывки ионообменника водой проводится десорбция лизина 0,5-5,0%-ной аммиачной водой, получаемый раствор содержит 80-90% лизина от сорбированного количества. Элюат упаривается в вакууме при 60 °С до 30-50%-ного содержания аминокислоты. При помощи соляной кислоты устанавливают pH 4,9, раствор упаривают и кристаллизуют аминокислоту при охлаждении до 12-14 °С. Кристаллы монохлоргидрата лизина отделяют на нутч-фильтре, промывают этиловым спиртом и высушивают при 60 °С. Форма готового продукта: L-лизин монохлоргидрат, содержание основного вещества 97-98%, влажность 0,5%, зольность до 0,3%, температура плавления 210 °С. Выход лизина на стадии выделения 76-78%.
Для животноводства Институтом биохимии им. А. Баха АН СССР совместно с Институтом микробиологии им. А. Кирхенштейна АН Латвийской ССР на Ливанском опытном биохимическом заводе было налажено производство кормового концентрата лизина - ККЛ (рис. 5.2).
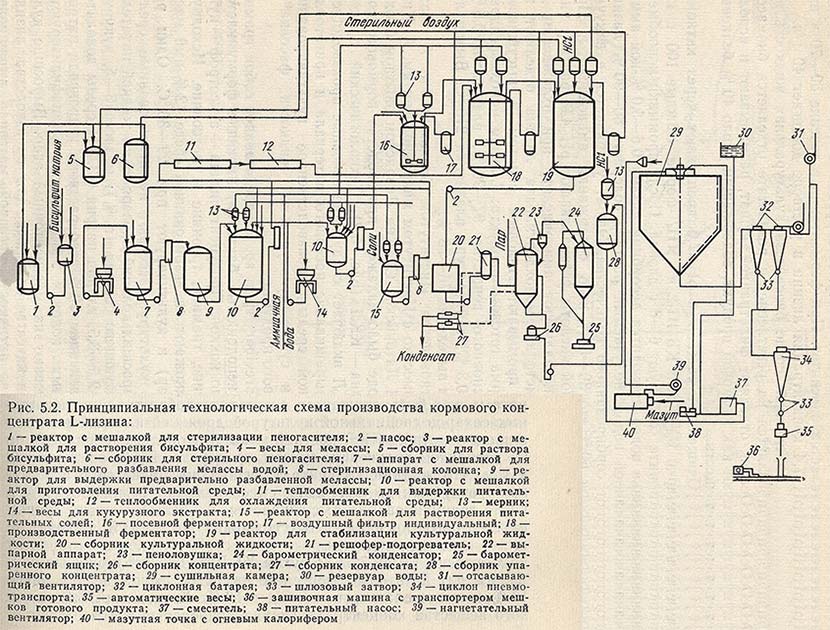
Технологический процесс получения ККЛ включает следующие основные производственные стадии: выращивание посевного материала и приготовление питательной среды, производственное культивирование, выпаривание и сушка культуральной жидкости, фасовка и упаковка готового продукта.
Культура Brevibacterium sp. 22 представляет собой грамположительные неподвижные бактерии, имеющие форму клеток от овальных до кокков. Продуцент лизина - ауксотроф - нуждается в биотине, тиамине, треонине и метионине. Исходную культуру размножают на агаризованной среде (2%-ный мясопептонный агар) и культивируют при 29-30 °С. Один раз в два месяца культуру рассевают на агаризованные среды. Активность выросших колоний проверяют на жидкой питательной среде следующего состава (в %): меласса 3-5, кукурузный экстракт 2,5-3,5, хлорид натрия 0,3. pH среды доводится до 7-7,2 добавлением 20%-ного раствора гидроксида натрия. Часть активных колоний высушивают (лиофилизируют), а часть пересевают на мясопептонный агар. Выросшие культуры служат в качестве исходных для производства лизина. Проверенный на активность посевной материал выращивают на мелассно-кукурузной среде в колбах (на 750 мл) в течение 24 ч при pH 6,9-7,0 и температуре 29-30 °С. Посевной материал характеризуется титром клеток 2,0*10 в 8 степени в 1 мл среды. Для выращивания культуры-продуцента в посевном аппарате готовят среду следующего состава (в %):

Среду стерилизуют 1 ч при температуре 126 °С. В посевной ферментатор на 250 л вносят 3-5% (от объема среды) посевного материала. Коэффициент заполнения посевного аппарата равен 0,5. Культуру выращивают при 29-30 °С, непрерывной аэрации (1 объем на 1 объем среды в минуту) и перемешивании (мешалка турбинного типа, 300 об/мин) в течение 24 ч. Для пеногашения добавляют 0,3% (от объема среды) простерилизованного (в течение 1 ч при температуре 120 °С) кашалотового жира.
Производственное культивирование продуцента. Производственное культивирование осуществляется в ферментаторах вместимостью 50 и 100 м3. До засева посевным материалом ферментатор промывается и стерилизуется в течение 1 ч при 0,1 МПа. Культура выращивается на среде следующего состава (в %):

Питательная среда стерилизуется при температуре 130-132 °С в течение 10-15 мин. После охлаждения до 30-32 °С среду подают в ферментатор. По стерильной посевной линии в ферментатор поступает посевной материал в количестве 5-6% от объема питательной среды. Коэффициент заполнения ферментатора составляет 0,75. Процесс ферментации продолжается 48-72 ч при температуре 29-30 °С, непрерывном перемешивании и аэрации среды стерильным нагретым воздухом (до 50 °С) из расчета 1 объем на 1 объем среды в минуту при избыточном давлении воздуха в ферментаторе 20-30 кГТа. Пеногашение осуществляется стерильным кашалотовым жиром, подаваемым из мерника пеногасителя. Общий расход пеногасителя составляет 0,5% от объема среды, причем 0,2% пеногасителя вносят в момент приготовления питательной среды.
Выпаривание и сушка культуральной жидкости. Стабилизированная 0,15%-ным бисульфитом натрия культуральная жидкость с pH 5,0-6,0 выпаривается на вакуум-выпарной установке. Начальная концентрация сухих веществ в жидкости, поступающей на выпарку, составляет 10-15%, конечная - около 40%. Упаренная культуральная жидкость высушивается нагретым воздухом на распылительной сушилке с дисковым распылителем при 300/90 °С.
Фасовка и упаковка готового продукта. Высушенный до остаточной влажности 4-8% ККЛ фасуется по 20 кг в крафт-мешки с полиэтиленовым вкладышем. При правильном соблюдении технологического режима выход лизина на стадии ферментации составляет 23-26% от содержания усвоенного сахара, а в культуральной жидкости накапливается до 20-25 г/л L-лизина. Общие потери лизина на стадии выпаривания и сушки не превышают 15%.
Сухой концентрат кормового лизина, получаемый высушиванием стабилизированной и сконцентрированной жидкости, имеет существенный недостаток. Он очень гигроскопичен и при хранении слеживается крупными комками. Гигроскопичность препарата может быть снижена в результате дображивания остаточных сахаров специальной культурой дрожжей или добавлением в процессе сушки ККЛ костной муки, бентонита, аэросила. Один из вариантов получения сухого препарата ККЛ заключается в том, что жидкий концентрат лизина смешивается с пшеничными отрубями до получения смеси влажностью около 70% и после гранулирования высушивается на конвективных сушилках. Готовый препарат ККЛ содержит до 7-10% лизина, он сыпуч и негигроскопичен.
Согласно ТУ 59-72-74 сухой ККЛ должен содержать монохлоргидрат L-лизина в пересчете на сухое вещество не менее 10% и иметь влажность не более 10%. Химический состав сухого вещества концентрата лизина приведен ниже.

Комбинированный, или энзиматический, способ производства лизина
Японская фирма «Тойо Рейон» («Торей») предложила в 1973 г. принципиально новый способ получения L-лизина, преимущество которого состоит в том, что конечный продукт отличается высокой концентрацией и чистотой. В ходе реакции не возникает опасности заражения посторонней микрофлорой. Отпадает необходимость разделения аминокислоты на оптические изомеры. Возможно также создание непрерывного процесса с использованием совместного действия иммобилизованных ферментов в одном реакторе.
Процесс состоит из стадий органического синтеза и ферментативного гидролиза. Исходным сырьем для получения этой аминокислоты служит циклогексан. В результате химических реакций из циклогексана получается циклический ангидрид лизина (DL-a-амино-е-капролактам), который на стадии ферментативного гидролиза превращается в L-лизин.
При этом производят разделение оптических изомеров, являющееся сложным и дорогостоящим процессом при синтезе не только L-лизина, но и других аминокислот. Разделение оптических изомеров DL-а-амино-е-капролактама осуществляется при использовании микробных ферментов. При этом происходит асимметрический гидролиз с помощью гидролазы аминокапролактама с образованием L-лизина.
Схема получения L-лизина, разработанная фирмой «Торей», представлена ниже.
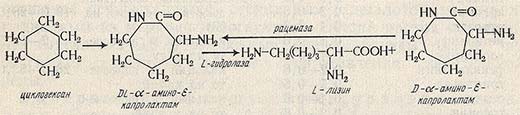
Помимо лизина в реакционной смеси содержится D-a-амино-e-капролактам, который с целью увеличения выхода L-лизина подвергают рацемизации. Для выявления преимущества рассматриваемого процесса необходимо, чтобы одновременно с гидролазой L-a-амино-е-капролактама действовал фермент, рацемизующий D-a-амино-е-капролактам. Только совместное действие этих двух ферментов обеспечивает превращение DL-а-амино-е-капролактама в L-лизин почти со 100%-ным выходом. Содержание аминокислоты в реакционной смеси может достигать 200 г/л.
Установлено, что способностью синтезировать гидролазу L-a-амино-е-капролактама обладают дрожжи родов Cryptococcus, Candida, Trichosporon. Активаторами этого фермента служат ионы Mn2+, Mg2+, Zn2+, ингибитором - этилендиаминтетрауксусная кислота. Фермент рацемазу а-амино-е-капролактама получают при выращивании бактерий, относящихся к родам Achromobacter, Flavobacterium и др. Ингибирующее действие на этот фермент оказывает гидроксиламин. Принцип действия рацемазы а-амино-е-капролактама, по-видимому, такой же, как и рацемаз других аминокислот, требующих в качестве кофактора пиридоксаль-5-фосфат.
Совместное действие двух ферментов на субстрат достигается введением в водный раствор DL-аминокапролактама определенного количества биомассы или сухих клеток дрожжей, обладающих гидролазной активностью, и бактерий с рацемазной активностью аминокапролактама. В результате реакции DL-аминокапролактам количественно переходит в L-лизин. Оптическая чистота получаемой аминокислоты более 99%.
Новый способ получения лизина интересен еще и тем, что L-гидролазу и рацемазу аминокапролактама можно получать, в иммобилизованной форме, что дает большие преимущества по сравнению с использованием растворимых ферментов. Для этого раствор DL-аминокапролактама, полученный химическим способом, пропускают через колонку, содержащую два иммобилизованных фермента микробного происхождения. Первый из них гидролизует амидную связь в L-аминокапролактаме, не затрагивая D-изомера, второй фермент - рацемаза - с высокой скоростью превращает D-изомер в рацемат. При правильно подобранных условиях и режиме работы колонки единственным продуктом процесса является L-лизин, его выход составляет 95%.